






最新资讯



-
电话: 400-113-6988 邮箱: dongfangxicao@163.com 地址: 深圳市南山区粤海街道高新中三道9号环球数码大厦19楼
↓ 微信扫码识别关注我们 ↓ 
Nrf2介导运动诱导的小鼠中线粒体生物发生和抗氧化反应
发表于:2020-06-01 作者:admin 来源:本站 点击量:11821
Nuclear factor erythroid-derived 2-like 2 (NFE2L2, Nrf2) mediates exercise-induced mitochondrial biogenesis and the anti-oxidant response in mice
Nrf2介导运动诱导的小鼠中线粒体生物发生和抗氧化反应
Merry T L , Ristow M . Journal of Physiology, 2016, 594(18):5195.
Nrf2介导运动诱导的小鼠中线粒体生物发生和抗氧化反应
Merry T L , Ristow M . Journal of Physiology, 2016, 594(18):5195.
关键点
·活性氧(ROS)和一氧化氮(NO)调节运动诱导的核因子红细胞2相关因子2(NFE2L2,Nrf2)在骨骼肌中的表达。
·NFE2L2(Nrf2)是急性运动诱导的骨骼肌线粒体生物发生基因增加所必需的,这些基因有:如核呼吸因子1(NRF-1)和线粒体转录因子A,以及抗氧化基因,如超氧化物歧化酶(SOD)1,SOD2和过氧化氢酶。
·NFE2L2(Nrf2)表达受损的小鼠运动训练后的运动表现、能量消耗、线粒体体积和抗氧化活性都会降低。
·在肌肉细胞中,ROS和NO可通过NFE2L2(Nrf2) / NRF-1依赖性途径调节线粒体生物发生。
摘要
有规律的运动可以诱导骨骼肌的适应性,包括线粒体的生物发生和增强抗氧化储备。这些适应和其他因素至少在一定程度上是身体活动个体健康状况改善的原因。活动期间产生活性氧(ROS)和一氧化氮(NO),可能介导对骨骼肌运动的适应性反应。然而,它们的作用机制尚不清楚。在本研究中,我们旨在确定氧化还原敏感转录因子核因子红细胞衍生2样2(NFE2L2/ Nrf2)在急性运动和训练诱导的线粒体生物发生和抗氧化反应中的作用。我们报道ROS和NO调节小鼠骨骼肌和肌肉细胞中急性运动诱导的NFE2L2(Nrf2)表达,并且NFE2L2的缺乏阻止了正常急性跑步机运动诱导的线粒体生物发生标志物mRNA的增加,如核呼吸因子1(NRF-1)和小鼠腓肠肌中的线粒体转录因子A(mtTFA)和抗氧化剂超氧化物歧化酶(SOD)1和2,以及过氧化氢酶。此外,经过5周的跑步机运动训练后,与野生型同窝仔相比,缺乏NFE2L2的小鼠运动能力和全身能量消耗以及骨骼肌线粒体质量和SOD活性均降低。在C2C12成肌细胞中,外源性H2O2(ROS)和二乙烯三胺/ NO加合物(NO供体)的急性处理诱导了mtTFA的增加,这通过小干扰RNA和NFE2L2或NRF-1的短发夹RNA敲低来阻止。我们的研究结果表明,在运动过程中,ROS和NO可通过NFE2L2发挥作用,在功能上调节骨骼肌线粒体生物合成和抗氧化防御基因的表达。
缩写: AICAR,5-氨基咪唑-4-甲酰胺核糖核苷酸; AMPK,5腺苷单磷酸激活的蛋白激酶; COX4,细胞色素c氧化酶亚基4; Deta / NO,二亚乙基三胺/一氧化氮加合物; GSH,还原型谷胱甘肽; GSR,谷胱甘肽还原酶; GSSG,氧化型谷胱甘肽; GST,谷胱甘肽S-转移酶; H2O2,过氧化氢; KO,敲除; L-NAME,NG-硝基-L-精氨酸; MAPK,丝裂原活化蛋白激酶; mtTFA,线粒体转录因子A; NAC,N-乙酰半胱氨酸; NFE2L2/Nrf2,核因子红细胞2相关因子2; NF-κB,活化B细胞核因子κ-轻链增强子; NO,一氧化氮; NRF,核呼吸因子; PGC1α,过氧化物酶体增殖物激活受体γ共激活因子1α; ROS,活性氧; shRNA,短发夹RNA; siRNA,小干扰RNA; SOD,超氧化物歧化酶; WT,野生型。
1. 引言
有规律的定期运动可以降低疾病发生率并延长预期寿命(Warburton等,2006)。骨骼肌对急性运动应激特别敏感,运动训练引起适应性变化,如抗氧化能力、线粒体体积和胰岛素信号中间体的增加,这改善肌肉功能,防止发育代谢紊乱(Egan&Zierath,2013)。运动可促进骨骼肌适应性变化的机制之一是通过急剧升高氧化应激和亚硝化应激来实现(Gomez-Cabrera等,2015)。
在运动或收缩过程中,骨骼肌产生活性氧(ROS)和一氧化氮(NO) (Powers & Jackson, 2008)。NO和ROS均可促进线粒体生物合成(Suzuki等,1998; Nisoli等,2003; Piantadosi&Suliman,2006)。在一些研究中,已经显示通过补充抗氧化剂来预防运动期间ROS的增加来减弱急性运动和运动训练诱导线粒体生物发生标志物和抗氧化剂反应的增加(Gomez-Cabrera等人2005; Gomez-Cabrera等人2008; Ristow等人2009; Strobel等人2011; Wadley等人2013; Paulsen等人,2014a),但在其他研究中没有这样的发现(Yfanti等人2010; Higashida等人2011; Wadley等人2013)。然而,ROS和NO协调运动应激适应性反应的机制尚不清楚。
转录因子过氧化物酶体增殖活化受体γ共激活因子1α(PGC1α)被视为运动诱导的线粒体生物发生和抗氧化反应的主要调节因子(Lin等人,2005)。PGC1α在运动后增加,并共激活核呼吸因子NRF-1和NRF-2,其激活线粒体转录因子A(mtTFA)以促进线粒体复制(Bassel-Duby&Olson,2006)。实际上,NO和ROS可以增加骨骼肌中PGC1α,NRF1和mtTFA的表达,并且抗氧化剂补充可以减少运动诱导的PGC1α水平的增加(Nisoli等人2003; Gomez-Cabrera等人2005; Ristow等人,2009)。这些观察结果表明ROS通过PGC1α起作用以调节骨骼肌中运动诱导的适应性反应。然而,PGC1α不是正常运动诱导的线粒体生物发生和抗氧化剂表达标志物增加所必需的(Leick等人,2008),表明替代机制也参与调节骨骼肌适应运动。
核因子红细胞衍生2样2(NFE2L2;也经常称为Nrf2)是ROS和NO敏感的转录因子(Kensler等人,2007)。细胞暴露于氧化应激或亚硝化应激导致NFE2L2从细胞质(在细胞质中NFE2L2被Keap1锚定来降解)转移到细胞核,在细胞核中它与抗氧化反应元件结合以启动保护细胞免受细胞毒性和氧化损伤的防御(Kensler等,2007)。NFE2L2的主要功能是协调抗氧化剂对应激的反应,因此,NFE2L2切除可以防止在最后一次非常激烈的短期跑步训练(2周)后,老年(> 23个月)小鼠骨骼肌抗氧化基因mRNA水平的立即增加 (Narasimhan等,2014)。此外,NFE2L2是心肌中一氧化碳应激(Piantadosi等人,2008)和肝脏炎症(Piantadosi等人,2011)诱导的线粒体生物发生所必需的。
因为NFE2L2可以调节线粒体生物发生和对急性氧化和亚硝化应激的抗氧化反应,我们确定骨骼肌NFE2L2是否被急性运动激活以及NFE2L2是否调节急性和运动训练诱导的线粒体生物发生和抗氧化剂响应的增加。我们报道NO和ROS均有助于在急性运动后增加骨骼肌NFE2L2的mRNA,并且NFE2L2表达是线粒体生物发生标志物正常增加和急性运动以及运动训练后骨骼肌抗氧化反应所必需的。此外,我们的数据表明,ROS(H2O2)和NO诱导的线粒体生物发生标记物(mRNA)的增加依赖于NFE2L2和NRF1,而只有NO介导的线粒体生物发生基因表达需要PGC1α。
2. 方法
2.1 动物伦理
本研究中描述的所有程序均经瑞士苏黎世兽医办公室(许可证号ZH245 / 14)批准,并按照苏黎世联邦理工学院和瑞士国家动物福利指南进行。研究人员了解生理学期刊运作的伦理原则,这项工作符合动物伦理检查表。
2.2 小鼠繁殖和住房条件
小鼠被养在一个温度控制的设施中,保持12:12小时的光/暗循环,并且可以随意食用不含维生素E、C(S8022-S005;SSNIFF Spezialdiaten GmbH,Soest,Germany)和水的饮食。先前已经描述了NFE2L2-/-(也称为Nrf2-/-)小鼠(Chan等人,1994),是购于Jackson 实验室(Bar Harbor,ME,USA)并在C57Bl / 6背景下内部繁殖。雄性(约25-30g,15-30周龄)NFE2L2-/-小鼠和NEF2L2 + / +同窝小鼠,分别称为NFE2L2敲除(KO)和野生型(WT),并且C57Bl / 6小鼠应用于所有的实验。所有小鼠均经颈椎脱位人工处死。
2.3 运动协议和代谢笼
所有运动小鼠在初始运动试验前5天以上分别接受至少两次跑步机(Panlab/Harvard Instrument,Holliston,MA,USA)熟悉训练。熟悉训练包括以0-9 米/ 粉种的速度于跑步机跑步5分钟,倾斜角度为0°。通过位于跑步机带末端的冲击网格提供温和的电刺激作为动力。有氧能力是通过增量跑步机测试至力竭来测定的。将跑步机设定为10°倾斜,并且每2分钟将速度增加2米/分钟,直到小鼠在不尝试继续跑步的情况下在冲击网格上度过>5秒。为了测试耐力能力,将跑步机设定为倾斜15°,并且起始速度为5米/分钟,其以0.17米/分钟的速率连续增加(倾斜),直到小鼠在不尝试继续跑步的情况下在冲击网格上度过>5秒。
运动训练包括30-60分钟的跑步机运动,以10-15米/分钟的速度运行,10°倾斜,4-5天/周,持续4-6周。急性运动方案包括1小时的跑步机运动训练,以12米/分钟的速度运行,倾斜10°。在指定的时间点进行急性运动后,以及在运动训练小鼠的最后一次运动至少36小时后,收集组织并在液氮中快速冷冻。
在24-48小时的适应期后,使用PhenoMaster(TSE 系统,Bad Homburg,Germany)开路量热系统测量超过48小时(两个光/暗循环)的耗氧量和动态活动。
2.4 抗体和试剂
兔NFE2L2抗体购自Santa Cruz Biotechnology (Dallas, TX, 美国), NRF-1购自Cell Signaling Technology (Beverly, MA, 美国)。5-氨基咪唑-4-甲酰胺核糖核苷酸(AICAR)购自Adipogen AG(Liestal,瑞士)。除非另有说明,否则所有其他试剂均购自Sigma-Aldrich Chemicals(St Louis,MO,美国)。
2.5 N -乙酰半胱氨酸(NAC)和L-NG-硝基精氨酸甲酯(L-NAME)处理小鼠
对于涉及抑制NO合酶和清除ROS小鼠的实验,实验前3天在小鼠饮用水中分别加入L-NAME(1 g/l)(Wadley&McConell,2007)或NAC(10 g/l)(Chen等人,2010)(Cobley等,2015)。
2.6 实时PCR
使用Trizol试剂(Invitrogen,Carlsbad,CA,美国)从细胞或冷冻肌肉样品中提取RNA,并使用高容量cDNA逆转录试剂盒(Applied Biosystems,Foster City,CA,美国)逆转录mRNA。为了定量mtDNA,通过标准蛋白酶K和苯酚 - 氯仿提取分离总DNA。使用SYBR green select master mix试剂盒(Applied Biosystems)在ViiATM 7实时PCR系统(Applied Biosystems)上进行定量实时PCR。反应一式三份进行,并使用ΔΔCt方法以18S核糖体RNA作为内部对照实现相对定量。使用的引物序列列于表1中。
表1. 小鼠定量PCR引物序列

2.7 细胞培养
C2C12成肌细胞在含有5.5mM D-葡萄糖和20%胎牛血清的DME培养基中于37℃在5%CO2比21%O2中生长。通过在抗生素选择(10天,1μg/ ml 嘌呤霉素)之前使用脂质体(Lipofectamine)2000(Invitrogen)转染合并的shRNA克隆(TRCN0000012128,TRCN0000012129,TRCN00000121; Broad Institute RNAi consortium,Cambridge,MA,美国)来实现NFE2L2的短发夹RNA(shRNA)敲低。通过使用Lipofectamine 2000用指定浓度的相应特异性小干扰RNA(siRNA)(Santa Cruz Biotechnology)转染来实现NRF-1和PGC1α基因表达的瞬时敲低,并在转染后24小时收集细胞。用AICAR(1mM),二亚乙基三胺/一氧化氮加合物(Deta / NO)(100μM)或H2O2(5μM)对C2C12细胞进行长时间的间歇处理,包括将化合物施加5小时/天,持续5天。剂量和治疗时间基于先前研究中研究永生化肌肉细胞中线粒体生物发生的浓度(McConnell等人2010年;Lira等人2010a),以及初步研究,这些研究获得了在不导致任何细胞死亡的情况下提高NFE2l2表达所需的最佳剂量(数据未显示)。
2.8 免疫印迹
基本上如先前所述(Merry等人,2014)进行免疫印迹操作。简而言之,将细胞从培养皿中分离出来,或者用电动手持匀浆机将冰冻的小鼠组织样本匀浆到10-20体积的RIPA冰裂解缓冲液中(50mM Hepes,pH 7.4,1%,v / v Triton X-100,1%v / v脱氧胆酸钠,0.1%v / v SDS,150mM NaCl,10%v / v甘油,1.5mM MgCl2, 1mM EGTA,50mM氟化钠,蛋白质抑制剂混合物(罗氏,巴塞尔,瑞士),1mM苯基甲基磺酰氟,1mM钒酸钠),在冰上孵育20分钟,并在4℃下以20,000g离心60分钟。通过SDS-PAGE分离上清液,并通过标准程序进行免疫印迹处理。
2.9 SOD和柠檬酸合酶活性测定
将冷冻肌肉与50mm磷酸盐缓冲液+1mMEDTA一起置于氮气冷冻砂浆中研磨,并进行超声处理。裂解液在12,000 g和4℃下离心15分钟。如上所述,将上清液用于随后的超氧化物歧化酶测量(Weimeret al.2014)。通过检测在412nm波长下5,5-二硫代双-2-硝基苯甲酸酯的增加,在上清液中测量柠檬酸合酶活性(Srere,1969)。
2.10 谷胱甘肽和蛋白质羰基化
使用BIOXYTECH GSH / GSSG-412测定试剂盒(Oxis International,Inc.,Tampa,FL,USA)测量全血中总的谷胱甘肽和氧化型谷胱甘肽水平,并根据制造商的说明书测定GSH:GSSG比率。根据制造商的说明书使用OxyBlot蛋白质氧化检测试剂盒(Merck Millipore,Billerica,MA,USA)监测蛋白质羰基化。
2.11 统计分析
所有数据均以平均值±SEM表示。使用非配对双尾Student’s t 检验和单向、双向ANOVA来确定统计学显着性,所述ANOVA具有所示的Fisher最不显着差异事后归因分析。P <0.05被认为具有统计学意义。
3. 结果
3.1 通过运动增加骨骼肌NFE2L2的表达
运动增加骨骼肌产生ROS和NO (Powers & Jackson, 2008),转录因子NFE2L2对细胞ROS和NO水平的变化敏感(Kensler等,2007)。因此,我们首先确定运动是否调节NFE2L2在小鼠骨骼肌中的表达。急性跑步机运动30分钟后,骨骼肌NFE2l2及其一些已知下游靶点(谷胱甘肽S-转移酶(GST)和谷胱甘肽还原酶(GSR))的mRNA增加了3倍以上(图1a)。这种增加是短暂的,并且在运动后2或16小时与休息的肌肉没有差别(图1B)。此外,NFE2L2,GST和GSR 的mRNA在运动训练的小鼠的骨骼肌中升高(图1C)。尽管测试了几种市售抗体,但我们无法鉴定出在组织样品中特异性检测到NFE2L2的抗体(图1D); 然而,我们通过测定骨骼肌、肝脏和心脏中的NFE2L2 mRNA水平,证实NFE2L2 KO小鼠不表达NEF2L2(图1E)。图1D中显示的印迹是使用NFE2L2抗体#12721(Cell Signaling Technologies)获得的,但是代表在小鼠肌肉,心脏和肝脏样本中测试的抗体#8882(Cell Signaling Technologies)和H300和C20(Santa Cruz Biotechnology)。重要的是要承认,尽管NFE2L2具有65-70kDa的预测分子量,但认为其在95-110kDa处被检测到(Lau等人,2013)。因此,我们无法确认NFE2L2 mRNA表达导致组织样品中蛋白质水平增加。然而,由于NFE2L2主要作为转录因子,因此其自身和其他靶标(GST和GSR)的mRNA增加证实它受运动调节。
在确定急性运动和运动训练增加骨骼肌NFE2L2 mRNA后,我们接下来研究了可能在运动过程中诱导NFE2L2表达的刺激因素。使用模拟运动的化合物处理C2C12成肌细胞,包括5 '腺苷单磷酸活化蛋白激酶(AMPK)激活剂(AICAR)、过氧化氢(H2O2)和NO供体Deta/NO增加NFE2L2蛋白表达(图1F)。为了确定ROS和NO是否在体内调节运动诱导的NFE2L2 mRNA的增加,在急性运动之前,我们用抗氧化剂NAC或NO合酶抑制剂L-NAME来处理小鼠。NAC将血液还原型谷胱甘肽(GSH)增加至氧化型谷胱甘肽(GSSG)的比率,表明抗氧化作用和全身氧化应激减少(图1I)。运动诱导的骨骼肌NFE2L2 mRNA的增加被L-NAME和NAC减弱,NAC也减弱了运动诱导的NFE2L2靶GST和GSR mRNA的增加(图1G和H)。
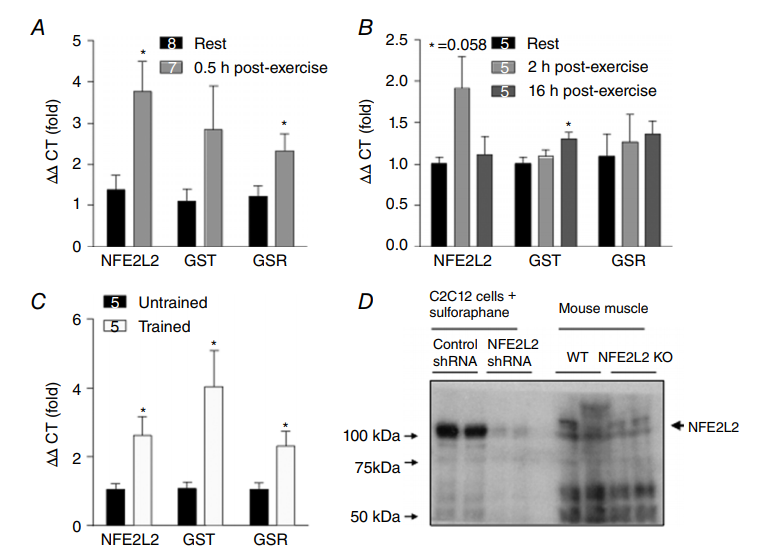
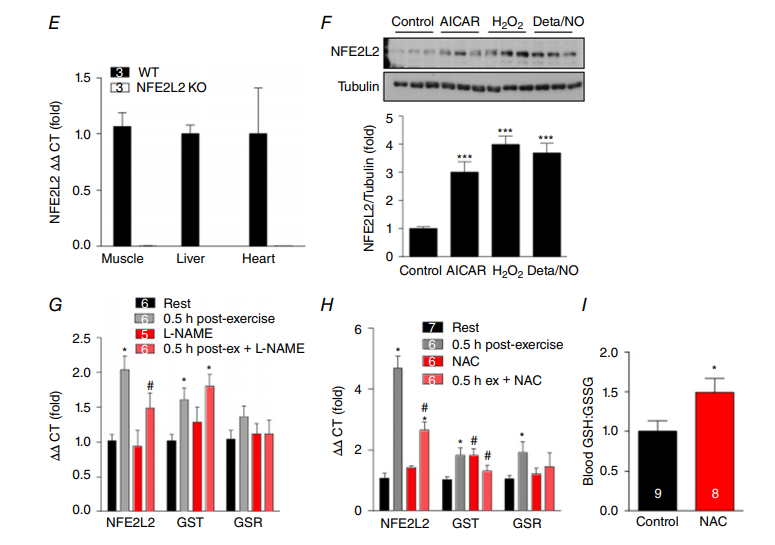
图1. 运动后骨骼肌中NFE2L2的表达
小鼠骨骼肌(腓肠肌)NFE2L2和NFE2L2靶基因(GST和GSR) mRNA在急性(1小时)一轮跑步机运动(A和B)和6周跑步机运动训练(C)30分钟后升高。代表性印迹显示测试的NFE2L2抗体细胞在裂解物中检测到报道的NFE2L2大小(95-110kDa)的条带,而在NFE2L2 shRNA处理的细胞没有发现;然而,这些抗体无法在组织裂解液中检测到正确分子量的条带,而NFE2L2 KO小鼠的裂解液中则没有这种条带(D)。NFE2L2 KO小鼠在骨骼肌,肝脏或心脏中不表达NFE2L2 mRNA(E)。用AICAR(1mM)和H2O2(50μM)或Deta / NO(100μM)急性(5小时)处理C2C12成肌细胞增加NFE2L2蛋白表达(F)。用NO合成酶抑制剂(G)或抗氧化剂(NAC)(H)治疗小鼠三天减弱了急性(1小时)运动诱导的骨骼肌(腓肠肌)NFE2L2、GST和GSR mRNA表达的增加和(I)增加血液GSH:GSSG比率。结果显示为平均值±SE(细胞培养实验的每组n = 5-7;小鼠实验的n分别显示)。 使用双尾Student’s t检验或具有Fisher最不显着差异事后归因分析的单向ANOVA确定显着性。与休息/对照相比,分别*P<0.05和***P<0.001。与相同条件下未处理组相比,#P<0.05。 [彩图在wileyonlinelibrary.com上查看]
3.2 NEF2L2是运动训练诱导的线粒体生物发生标志物所必需的
ROS和NO参与调控运动诱导的线粒体生物发生(GomezCabrera et al. 2005;Gomez-Cabrera等,2008);而NFE2L2是心肌和肝脏应激刺激线粒体生物发生所必需的(Piantadosi et al. 2008;Piantadosi等人,2011)。因此,我们研究了NFE2L2是否是运动诱导的骨骼肌线粒体生物发生所必需的。运动训练增加了WT但不是NFE2L2 KO小鼠的骨骼肌中的mtDNA(线粒体DNA)和柠檬酸合酶活性(线粒体体积标记物)(图2A和B)。与此相一致,与相同基因型的未训练小鼠相比,经训练的WT而不是未训练的NFE2L2 KO小鼠的骨骼肌中的线粒体生物发生相关基因mRNA增加(图2C)。然而,由于个体差异很大,我们无法检测到PGC1α,mtTFA或细胞色素c氧化酶亚基4(COX4)的骨骼肌蛋白水平在训练或基因型上有差异(数据未显示)。
与线粒体生物发生数据一致,运动训练提高了WT小鼠的有氧和耐力,但没有提高NFE2L2 KO小鼠的有氧和耐力(图2D和E)。尽管对WT或NFE2L2 KO小鼠体重没有训练效果(图2F),训练的WT小鼠的全身能量消耗大于训练的NFE2L2小鼠的全身能量消耗(图2G),在未训练的小鼠中并没发现这样的效果并且不是活动水平差异的结果(数据未显示)(图2H)。
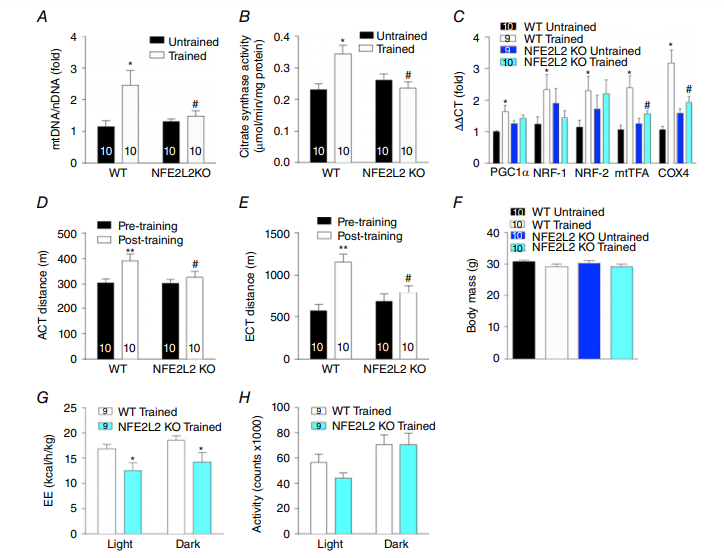
图2. NFE2L2是运动训练诱导的线粒体生物发生所必需的
在跑步机运动训练(训练)或正常久坐行为(未经训练)6周后,在WT和NFE2L2 KO小鼠的骨骼肌(腓肠肌)中测量mtDNA(A),柠檬酸合酶活性(B)和线粒体生物发生相关基因mRNA(C)。经过四周跑步训练的NRFE2L2 KO小鼠的运动表现(D和E)和能量消耗(G)均低于经过训练的WT小鼠。WT和NFE2L2 KO小鼠具有相似的体重和活动水平(F和H)。结果显示为平均值±SE(n分别显示)。使用具有Fisher最不显着差异事后归因分析的单向ANOVA确定显着性。与相同基因型的未训练或预训练小鼠相比,* P <0.05 ** P <0.001。与同条件下的WT相比,#P <0.05。 EE,能源消耗。 [彩图可在wileyonlinelibrary.com上查看]
3.3 ROS和NO通过NFE2L2和NRF-1起作用以诱导线粒体生物发生
为了进一步研究NFE2L2在运动诱导的线粒体生物发生中的作用,我们评估了急性运动对WT和NFE2L2 KO小鼠中线粒体生物发生相关基因mRNA的影响(图3A)。在急性运动后,WT和NFE2L2 KO小鼠的骨骼肌中PGC1α和COX4 mRNA相似地增加,而NRF-1和mtTFA mRNA仅在WT小鼠的骨骼肌中增加(图3A)。
然后,我们利用C2C12成肌细胞模型来研究NFE2L2可能通过何种途径促进骨骼肌线粒体生物发生。用AMPK激活剂(AICAR)对对照组和NFE2L2蛋白(NFE2L2 shRNA)缺乏的细胞进行急性处理(图1D)。尽管AICAR处理增加了对照和NFE2L2缺陷细胞中PGC1α和mtTFA mRNA水平,但NFE2L2 shRNA阻止了AICAR诱导的NRF-1和COX4 mRNA水平的增加(图3B)。同样,NFE2L2 shRNA也减弱了AICAR诱导的mtDNA长时间间歇性增加(图3C)。为了确定NFE2L2在NO或ROS介导的线粒体生物发生中的作用,对照组和NFE2L2 shRNA细胞分别暴露于NO供体Deta/NO或H2O2的急性处理(图3D和E)或长时间间歇处理中(图3F)。在对照组中,急性Deta/NO或H2O2处理增加线粒体生物发生基因mRNA,而NFE2L2 shRNA细胞则没有。同样地,使用DeTa/NO或H2O2长时间间歇处理增加了对照组的mtDNA,但没有增加NFE2L2 shRNA细胞的mtDNA(图3F)。
因为NRF-1的启动子区域含有NFE2L2结合位点(Piantadosi&Suliman,2006),并且NFE2L2缺乏不影响急性运动或AICAR诱导的PGC1α表达(图3A和B),我们使用siRNA来确定是否NRF-1和PGC1α是NO和ROS介导的下游线粒体生物发生介质mtTFA mRNA的增加所必需的。通过NRF-1的siRNA敲低阻止了Deta / NO和H2O2诱导的mtTFA mRNA增加(图3G),并且PGC1α siRNA敲低阻止了Deta / NO诱导的但不抑制H2O2诱导的mtTFA增加(图3H)。
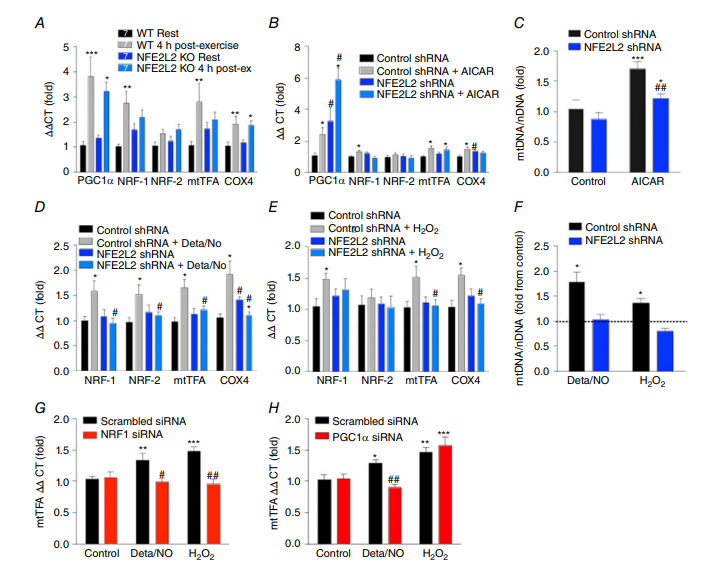
图3. NFE2L2是急性运动、NO和H2O2诱导线粒体生物合成相关基因mRNA增加所必需的
急性运动后4小时WT和NFE2L2 KO小鼠的骨骼肌(腓肠肌)中线粒体生物发生相关基因mRNA水平(A)。在用AICAR(1mM)(B),Deta / NO(100μM)(D)或H2O2(50μM)(E)处理5小时后,对照shRNA或NFE2L2shRNA C2C12成肌细胞中的线粒体生物发生相关基因mRNA表达。在用AICAR(1mM)(C),Deta / NO(100μM)或H 2 O 2(5μM)(F)处理5天(5小时/天)后,在对照shRNA或NFE2L2 shRNA C2C12成肌细胞中测定mtDNA。在对照组、NRF-1(100nM)(G)或PGC1α(10nM)(H) siRNA转染后24小时用Deta / NO(100μM)或H2O2(50μM)刺激C2C12成肌细胞持续5小时来评估mtTFA mRNA表达。结果显示为平均值±SE(对于细胞培养实验,每组n = 6-8;对于小鼠实验,n分别显示)。使用单因素方差分析和Fisher最不显着差异事后归因分析确定显着性。与相同基因型的休息或未处理小鼠或si/shRNA小鼠相比,* P <0.05,** P <0.01 *** P <0.001。与相同条件下的对照组si / shRNA小鼠相比,#P <0.05。 [彩图可在wileyonlinelibrary.com上查看]
3.4 ROS,NO和NFE2L2调节运动诱导的抗氧化反应
NO合成抑制剂L-NAME和抗氧化剂NAC阻止急性运动诱导的骨骼肌抗氧化基因超氧化物歧化酶(SOD)1,SOD2,过氧化氢酶,Prx1和Trx1 mRNA的增加(图4A和B)。虽然SOD1 mRNA水平升高,并且SOD2在未锻炼的NFE2L2骨骼肌中趋于升高,但急性运动并未进一步增加其水平,也没有增加过氧化氢酶mRNA(图4C)。相反,急性运动增加了WT小鼠骨骼肌中SOD1,SOD2和过氧化氢酶的mRNA水平(图4C)。在运动训练之后,SOD1,SOD2和过氧化氢酶mRNA在WT的骨骼肌中增加而在NFE2L2KO小鼠中没有增加(图4D),而Prx1的mRNA仅通过在NFE2L2KO小鼠的骨骼肌中的训练而增加。运动训练增加WT但不是NFE2L2 KO小鼠的骨骼肌SOD活性(图4E)。尽管具有较低的抗氧化水平,但NFE2L2 KO小鼠骨骼肌中的蛋白氧化(通过氧印迹蛋白羰基化实验评估)并没有升高,运动训练也没有改变蛋白氧化水平(图4F)。
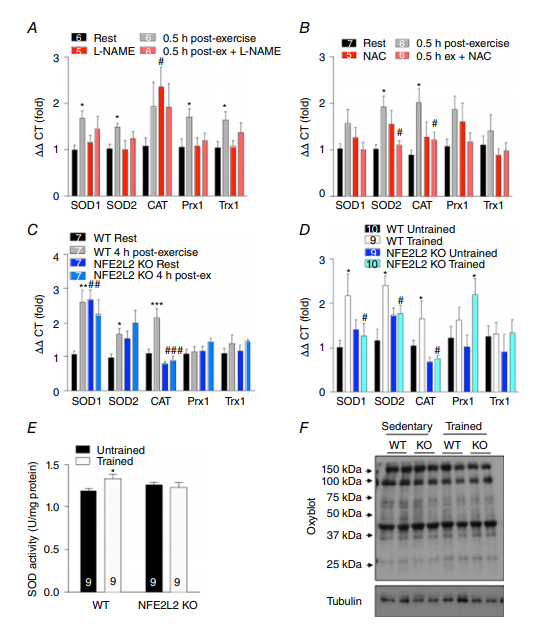
图4. NFE2L2,ROS和NO调节运动诱导的抗氧化反应
采用NO合酶抑制剂(L-NAME) (A)或抗氧化剂(NAC) (B)处理小鼠3天,在急性(1小时)平板运动后30分钟检测骨骼肌(腓肠肌)抗氧化剂基因mRNA的表达。急性平板运动(C)或6周平板运动训练(训练)或正常久坐行为(未经训练)后4 h, WT和NFE2L2 KO小鼠骨骼肌(腓肠肌)中抗氧化基因mRNA或SOD活性测定(D和E)。WT和NFE2L2 KO腓肠肌蛋白羰基化水平相似,且不受运动训练的影响(F)。结果显示为平均值±SE(n分别显示)。使用具有Fisher最不显着差异事后归因分析的单向ANOVA确定显着性。与休息或未经训练的相比,* P <0.05和*** P <0.001。 与未处理的或相同条件下的WT相比,#P <0.05。CAT,过氧化氢酶; Prx,过氧化物还原酶; Trx,氧碘氧还蛋白。 [彩图可在wileyonlinelibrary.com上查看]
4. 讨论
在本研究中,我们发现NO和ROS在急性运动应激后调节NFE2L2 mRNA水平,NFE2L2表达的中断损害了急性运动和运动训练诱导的骨骼肌线粒体生物发生标志物的增加和抗氧化反应。与这些发现一致,缺乏NFE2L2的运动训练小鼠降低了全身能量消耗和运动表现。本研究首次提供了运动诱导NO和ROS增加可能促进对运动压力适应机制的因果证据。
人类抗氧化剂补充(Gomez-Cabrera等,2008;Ristow等,2009;Paulsen等(2014a)和啮齿类动物模型(Gomez-Cabrera等,2005;Strobel等人,2011)已被证明能够减弱运动诱导的骨骼肌线粒体生物发生标志物的增加。本研究通过提供证据进一步证明,运动期间补充抗氧化剂会减弱NFE2L2的正常激活,并且NFE2L2是线粒体生物反应对运动的氧化还原敏感性的转录调节因子。运动激活骨骼肌中的AMPK和p38丝裂原活化蛋白激酶(MAPK),这些蛋白激酶被认为通过PGC1α起作用,触发转录因子NRF-1,NRF-2和mtTFA的反应从而诱导线粒体生物合成(Bassel-Duby&Olson,2006)。尽管抗氧化剂补充剂可以减轻急性运动引起的AMPK和p38 MAPK磷酸化和PGC1α表达的增加(Gomez-Cabrera等人,2005),这并不总是阻止运动训练导致线粒体生物发生的增加(Wadley等人2013)。此外,缺乏AMPKα2或PGC1α的小鼠对运动表现出正常的线粒体生物生成反应(Jorgensen等人2007年;Leick等人2008)。这表明其他氧化还原敏感途径可能参与调节响应运动的线粒体生物合成。
我们的研究发现,NFE2L2表达的中断会损害运动,NO和H2O2诱导的线粒体质量标记物(柠檬酸合酶和mtDNA)的增加得到了先前研究的支持,这些研究表明NFE2L2调节白藜芦醇、败血症和一氧化碳诱导的线粒体生物发生(Piantadosi et al. 2008;Piantadosiet。2011;Kim等,2014)。机制上,NRF-1启动子区含有NFE2L2结合位点(Piantadosiet al. 2011),表明NFE2L2激活可诱导NRF-1转录,随后NRF-1可与mtTFA相互作用,驱动线粒体生物发生。急性运动,NO和H2O2诱导的NRF-1和mtTFA的增加依赖于NFE2L2,并且在肌肉细胞中,通过破坏NRF-1来阻止NO和H2O2诱导的mtTFA增加。与我们的肌肉细胞数据一致,啮齿动物中的NO合酶抑制剂和抗氧化剂补充减少了急性运动后的mtTFA表达(Wadley&McConell,2007; Wadley等人,2013),并且在大鼠肝细胞瘤中,氧化应激增加了NRF -1与mtTFA的结合(Piantadosi&Suliman,2006)。这表明ROS和NO可以通过NFE2L2和NRF-1起作用以促进由mtTFA调节的线粒体生物发生。
为了支持这个概念:NFE2L2是线粒体生物发生信号级联中PGC1α下游途径的一部分,在缺乏NFE2L2的情况下,AICAR和急性运动均刺激PGC1αmRNA的正常增加。然而,有趣的是,PGC1α的siRNA敲低抑制了了NO但不抑制H2O2介导的mtTFA mRNA的增加。考虑到AICAR刺激NFE2L2表达和AICAR诱导的线粒体生物发生部分地被NFE2L2 shRNA敲低而抑制,我们推测在涉及PGC1α和NFE2L2促进线粒体生物发生的途径之间可能存在一定程度的合作。以前的研究表明NO通过涉及cGMP形成和PGC1α诱导的途径促进线粒体生物发生(Nisoli等,2003; Lira等,2010b)。虽然NO与Keap1半胱氨酸151相互作用以抑制NFE2L2的降解(McMahon等人,2010),从而促进NFE2L2表达,但尚不清楚这是否是运动诱导的NO促进NFE2L2与NRF-1相互作用的机制或是否涉及中间步骤,例如cGMP的形成。无论如何,由于H2O2似乎独立于PGC1α起作用,PGC1α和NFE2L2可以协同作用和/或是相互独立的观点需要进一步关注。
然而,有几个研究小组报告说,抗氧化剂补充剂不会影响正常的骨骼肌运动反应(Yfanti等人2010; Higashida等人2011; Wadley等人2013)。尽管可变的抗氧化剂补充剂(类型,持续时间和剂量)和运动训练(模式,持续时间,强度)可能是重要的促成因素(Merry&Ristow,2015),但这些支持运动线粒体生物发生途径的发现可能是多余的。实际上,我们表明ROS和NO都有助于急性运动后NFE2L2的激活,这可能表明,尽管清除一个可能有效地减少线粒体对运动信号的生物生成反应,但这种抑制作用已被克服,和/或替代途径正在上调,伴随着随后的发作导致正常的训练反应。此外,必须承认,在本研究中,我们使用了全身NFE2L2缺陷小鼠,尽管进一步的体内研究应该考虑这些小鼠的钝性运动反应是肌肉特异性作用的结果还是全身调节的结果。
NFE2L2调控从酵母、秀丽隐球菌到哺乳动物等生物体的基础抗氧化表达和对应激的抗氧化反应(Kensler et al. 2007;Nguyen等,2009)。最近,已显示NFE2L2在老年小鼠的骨骼肌(Narasimhan等人,2014)和年轻小鼠的心肌(Muthusamy等人,2012)中进行彻底运动后立即增加抗氧化剂mRNA表达。此外,我们还发现,ROS和NO都参与了NFE2L2在运动过程中的激活,且NFE2L2不仅是急性运动后抗氧化剂表达和抗氧化活性正常增加的原因,而且也是运动训练的结果。我们还报道SOD1和2的表达在基础(休息)状态下上调,并且与WT小鼠不同,在NFE2L2KO小鼠中通过运动训练增加过氧化物酶1的表达。我们还报道了在基础(休息)状态下SOD1和2的表达上调,与WT小鼠不同,在NFE2L2 KO小鼠中,过氧合蛋白1的表达随着运动训练的增加而增加。虽然这并没有转化为NFE2L2 KO小鼠基础SOD活性的增加,但这可能是过氧化氢酶和谷胱甘肽代谢相关基因表达减少的代偿性反应(数据未显示;Narasimhan etal . 2014),表明其他机制也调控骨骼肌抗氧化表达。一个可能的候选者是核因子κ-轻链增强子激活的B细胞(NF-κB),已知其参与抗氧化剂表达的转录调节,并且维生素C和别嘌呤醇补充抑制运动引起NF-κB表达的增加(Gomez-Cabrera等人2005; Gomez-Cabrera等人2008; Ji,2008)。
在一种被称为毒物兴奋效应(Southam & Ehrlich, 1943)的反应中,ROS和NO应激的急剧增加促进了适应性,保护细胞免受随后应激的潜在有害影响,并延长了秀丽线虫和黑腹果蝇的寿命(Ristow & Zarse, 2010)。切除线虫NFE2L2同源物(skin-1),消除了干预措施对寿命延长的作用,例如胰岛素/胰岛素样生长因子-1信号传导的损伤(Zarse等人,2012)、葡萄糖胺补充剂(Weimer等人, 2014)和糖酵解抑制(Schulz等人,2007)。重要的是,这些干预措施需要增加ROS来诱导适应性,包括增加线粒体呼吸和抗氧化表达,以延长寿命。因为定期的体育锻炼也会增加预期寿命(Warburton等人,2006),并且增强的线粒体呼吸和增加的内源性抗氧化剂表达可以预防代谢疾病(Koves等人,2008; Anderson等人,2009),因此很容易推测 NFE2L2可能参与运动的整个生物体健康促进作用。事实上,NFE2L2激活剂已被证明可以延长秀丽隐杆线虫和黑腹果蝇的寿命(Suckow&Suckow,2006; Lee等人,2010),并预防小鼠的代谢疾病(Yu 等,2011; He 等,2012)。然而,令人惊讶的是,NFE2L2的脂肪特异性和全身缺失也减少了遗传和饮食诱导的肥胖(Chartoumpekis等人2011; Xue等人2013)。这可能表明NFE2L2在调节代谢反应中的组织特异性作用,并表明与慢性缺失与急性激活相关的差异效应。无论如何,这些数据和以前发表的人类研究数据(Gomez-Cabrera等人2008; Ristow等人2009; Paulsen等人2014a; Paulsen等人2014b; Cobley等人2015)表明在正常生理条件下,特别是在运动过程中产生的ROS,是最佳适应和对未来细胞应激源的保护所必需的。这建议不要在人体中不必要地使用抗氧化剂。虽然只是推测,但我们的数据也可能表明,摄入激活NFE2L2的营养素可能对细胞功能有保护作用,类似于运动;然而,这还有待证实。这些结果与兴奋理论一致,其中ROS / NO应激的急剧增加启动细胞和可能的系统范围的适应性反应,其改善对预期应激的耐受性。
总之,我们的数据表明运动过程中产生的NO和ROS激活骨骼肌NFE2L2。此外,NFE2L2的表达是正常线粒体生物发生和对急性运动&运动训练的抗氧化转录反应所必需的。这些结果与毒物兴奋效应相一致,ROS/NO压力的急剧增加启动了细胞的适应性反应,可能还启动了整个系统的适应性反应,从而提高了对预期应激的耐受性。
本文由福山生物整理翻译,转载请注明出处。