






最新资讯



-
电话: 400-113-6988 邮箱: dongfangxicao@163.com 地址: 深圳市南山区粤海街道高新中三道9号环球数码大厦19楼
↓ 微信扫码识别关注我们 ↓ 
Nrf2介导氧化还原作用对运动的适应
发表于:2021-02-20 作者:admin 来源:本站 点击量:17574
原文:Done A J , Traustadóttir, Tinna. Nrf2 mediates redox adaptations to exercise[J]. Redox Biol, 2016, 10(C):191-199.
翻译:
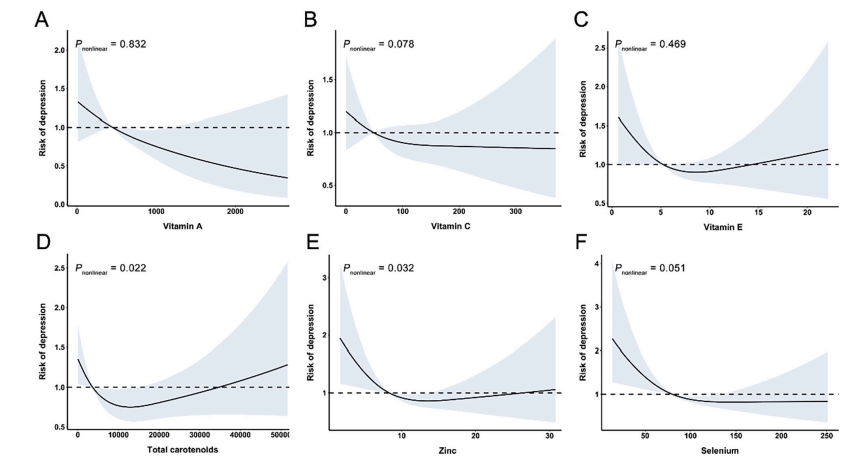
综述的主要目的是总结当前关于急性运动和定期运动对核因子红细胞样2相关因子2(Nrf2)活性和Nrf2信号下游靶点的影响的文献。Nrf2(在人体内由NFE2L2基因编码)是抗氧化剂防御的主要调节因子,是一种调节200多个细胞保护基因表达的转录因子。越来越多的证据表明,Nrf2信号在氧化应激如何介导运动的有益作用中起着关键作用。通过急性运动诱导的氧化应激偶发性增加刺激了Nrf2活化,与定期运动一样重复刺激时,会导致内源性抗氧化防御能力与总体更大的抗氧化应激破坏性作用能力的上调。运动造成的不同组织Nrf2激活证据可能是运动如何发挥其众所周知的系统效应的一个重要机制,这些系统效应不仅限于骨骼肌和心肌。此外,还有一些动物研究成果转化为人类的数据。
1、简介
随着分子生物学的发展及其在运动中的应用,人们对于急性运动诱导细胞信号导致多次运动后积累生理适应性的认识得到了极大提高。急性运动激活初级信使,如一磷酸腺苷(AMP)、钙和机械应激,以及随后的次级信使,包括AMP活化蛋白激酶(AMPK)、钙/钙调蛋白依赖性蛋白激酶(CAMK)和p38有丝分裂原活化蛋白激酶,导致mRNA转录的急性变化。急性运动刺激的另一个重要的初级信使是氧化还原平衡(或氧化还原电位)的暂时性变化,通过产生活性氧物质(ROS)进入更加氧化的状态。虽然ROS最初被认为具有损伤性,但现在人们对ROS在细胞信号传导、免疫功能调节、基因转录和凋亡中的重要作用有了更深入的了解。一个主要的例子是ROS在转录因子核因子红细胞样2相关因子2(Nrf2)激活中的作用,其是抗氧化剂酶和细胞抗应激的主要调节因子。啮齿动物研究的数据表明,靶向Nrf2活性以降低疾病风险的治疗潜力。靶向Nrf2敲除(Nrf2–/–)模型在所有组织中对毒性和氧化应激源表现出抵抗力明显受损,因此Nrf2–/–小鼠更易患慢性疾病。靶向Nrf2激活的药理学和遗传学(负调节子敲除)方法有效地改善氧化应激源造成的损害并恢复对慢性疾病的抵抗。Nrf2信号中断的情况下病理学的增加,以及随后在Nrf2 活化剂存在下的逆转已经在其他地方进行了综述。
越来越多的证据表明,Nrf2途径在氧化应激如何调节运动的有益作用中起着关键作用。急性运动诱导的氧化应激的偶发性增加会刺激Nrf2激活,与定期运动一样重复刺激时,这可能会导致内源性抗氧化防御的上调,以及总体上更大的能力以抵消氧化对核酸、蛋白质和脂质的损伤作用。本综述的主要目的是总结当前关于急性运动和定期运动或运动干预对Nrf2活性和Nrf2信号下游靶点影响的文献。我们还研究了支持或反对与运动结合使用抗氧化补充剂和植物营养素Nrf2活化剂的证据。由于干预效果在目标受损时更容易观察到,我们将着重研究衰老作为受损细胞信号的非疾病模型,其通常与年龄相关的病理学相协调。我们从Nrf2调节的简要概述开始。
2、 Nrf2:细胞防御的主调节子
转录因子Nrf2(在人类中由NFE2L2基因编码)是抗氧化防御的主调节子,调节超过200种保护氧化应激的细胞保护基因。Nrf2是碱性亮氨酸拉链(bZIP)转录因子家族的一员,在无应激条件下通过与胞质溶胶中的Kelch样红系细胞相关CNC同源关联蛋白1(Keap1)的同二聚体结合而被抑制[17]。Nrf2和Keap1之间的相互作用在物种间高度保守,表明其调节作用的重要[18]。在这种状态下,Keap1作为Nrf2降解的衔接蛋白发挥作用,其由Cul3/Rbx1介导,促进Nrf2泛素化和随后26s蛋白酶体对Nrf2的降解。Keap1/Cul3/Rbx1也作为一种额外的负调节因子存在于细胞核中[19]。
半胱氨酸残基被以一种独特的形式修饰来响应氧化或亲电刺激,不同的结构诱导物与Keap1上不同的半胱氨酸残基组合反应,从而产生相同的生物响应,特别是从Nrf2脱离并激活Nrf2-抗氧化反应元件(ARE)响应,或通过稳定Keap1-Nrf2复合物,减缓蛋白酶体对其的降解[20–23]。一旦从Keap1上脱离,Nrf2就转移到细胞核中,在那里它与小Maf蛋白异二聚体化,并结合顺式作用元件AREs,有效地激活II相解毒酶的转录(见图1)。
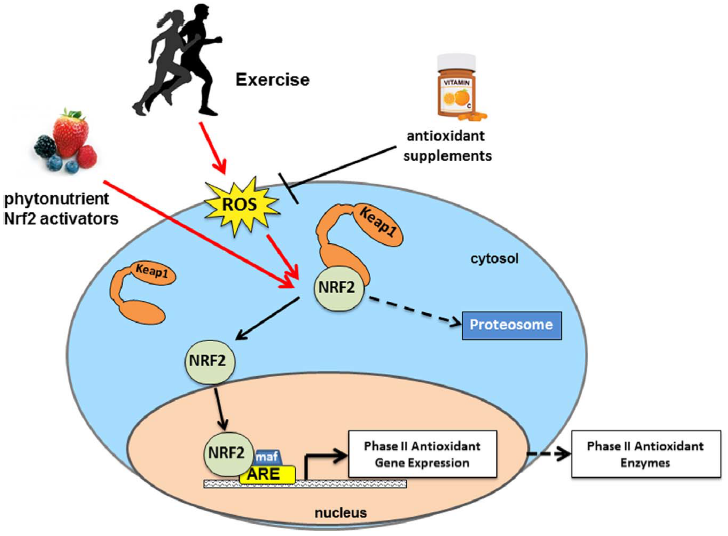
图1 Nrf2信号通路。Nrf2被运动诱导的ROS或植物营养素Nrf2激动剂激活。抗氧化补充剂抑制了运动诱导的ROS的信号,从而抑制下游的Nrf2信号传导。
在治疗潜力方面,这种半胱氨酸编码作为多种化合物和刺激物的有效Nrf2诱导物,之间相互独立,引起了人们极大的兴趣。例如,运动诱导的Nrf2的活性氧活化可能通过氧化这些半胱氨酸残基发生。同样,某些植物营养素被证明能够激活Nrf2,这个过程可能是通过改变半胱氨酸残基而发生的,而这些残基不同于运动中的靶点。如果事实上是这样的话,运动训练和Nrf2激动化合物之间可能存在协同效应。一些最新数据支持这一点,这些数据在本综述的第6节中给出。对于那些对Nrf2激活机制的更详细背景感兴趣的人,我们请读者参考这方面近期的一篇优秀评论[16]。
值得注意的是,在某些情况下,如癌症,Nrf2的激活和信号传导可能会产生有害影响。在正常情况下,Nrf2信号可以防止癌症,正如十字花科蔬菜中的植物营养素刺激Nrf2活化的效果一样。然而,在癌细胞中,Nrf2的激活可能赋予细胞抗应激能力,使细胞不易受到化疗治疗的影响。研究表明,在癌症肿瘤中,Nrf2被上调,在化疗期间可能需要使用Nrf2抑制剂提高治疗效果。
3、Nrf2信号对急性运动的反应
使用C2C12骨骼肌细胞进行的细胞培养为证明Nrf2被ROS激活的研究提供了证据,当抗氧化剂(如N-乙酰半胱氨酸)应用于培养基时,激活被抑制。Horie等人使用电脉冲刺激(EPS)模拟急性运动,并证明Nrf2表达与刺激强度和持续时间相关。此外,EPS刺激了Nrf2相关抗氧化基因的表达,当siRNA转染敲低细胞中的Nrf2时,表达减弱。其他研究表明,用H2O2处理肌管或大鼠心肌细胞后,Nrf2蛋白表达增加。
这些体外结果转化为体内结果主要在动物模型中完成,最近的一项研究是在临床研究。表1列出了这些研究。值得注意的是,早期的一些研究在其免疫印迹中针对一个非特异性的Nrf2(57 kDa)条带。尽管预测的Nrf2分子量为57 kDa,但一些研究小组已经确定90-110 kDa带是与生物相关的Nrf2靶点。
表1 急性运动诱导Nrf2信号
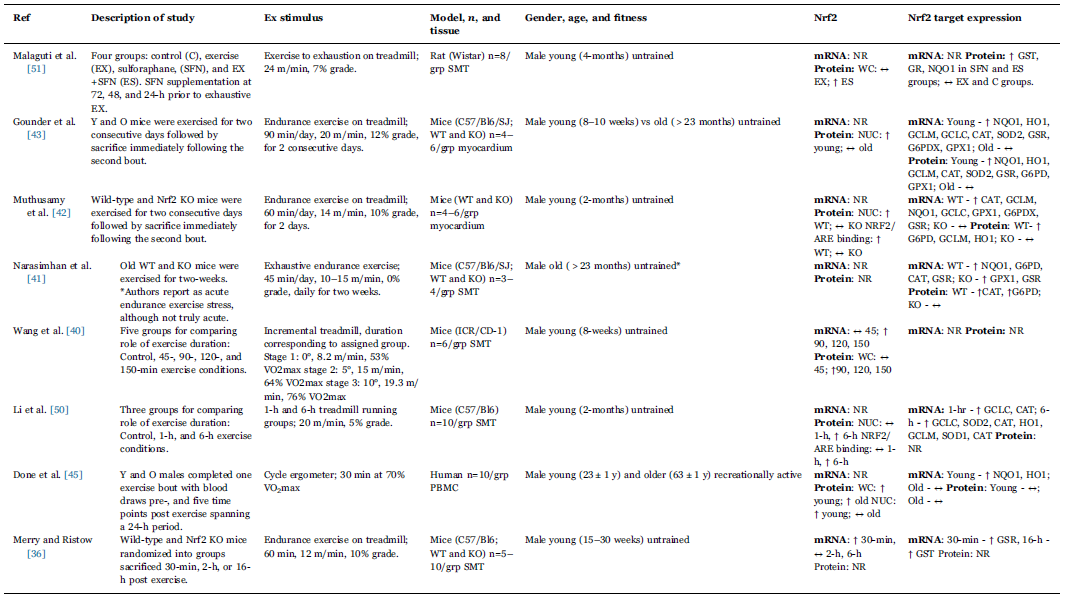
NR未报告或未测量,SMT骨骼肌组织,PBMC外周血单核细胞,NUC核,WC全细胞。
在野生型小鼠中,单次急性运动提高了骨骼肌中的Nrf2基因表达和其蛋白丰度,以及依赖Nrf2的II相酶。在另外两项测量运动对小鼠心肌中Nrf2信号传导影响的研究中,结果基本上与骨骼肌研究的数据一致。然而,这些研究使用了“急性运动应激”,其为连续两天60分钟的跑步机运动。虽然很明显时间不足以归类为运动训练,但众所周知,连续运动会导致mRNA表达的累积反应,可能需要观察蛋白质丰度的可测量变化[44]。然而,Muthusamy等人[42]提供了对运动反应的Nrf2信号的最全面分析,并比较了年轻野生型(WT)和Nrf2–/–小鼠。尽管两组的ROS都有相似的增加,但运动引起的核积累显著增加,并且Nrf2在WT小鼠心肌中结合,而在Nrf2–/–小鼠中没有观察到Nrf2活性的变化。在WT小鼠中几乎所有II相酶的基因表达都显著增加,唯一的例外是血红素加氧酶-1(HO-1)。II相抗氧化蛋白的表达变化不太一致,仅在HO-1、谷氨酸半胱氨酸连接酶(GCLM)和葡萄糖-6-磷酸脱氢酶(G6PD)中观察到变化。
关于运动诱导的人体Nrf2信号传导的数据仍非常有限,但最近的一项研究首次表明,在年轻和老年人的外周血单核细胞(PBMCs)中,急性运动增加了全细胞水平的Nrf2蛋白丰度[45]。此外,仅在年轻组中观察到Nrf2的核聚积,表明老化与Nrf2的核输入受损有关(见图2)。毫不奇怪,运动刺激了年轻人而不是老年人的HO-1和NAD(P)H醌脱氢酶1(NQO1)基因表达的增加。之前在小鼠心肌和大鼠肝脏中显示了与年龄相关的胞质溶胶和细胞核中Nrf2水平以及Nrf2/ARE结合能力的降低,并伴随响应急性运动的Nrf2依赖的抗氧化基因表达的减弱。
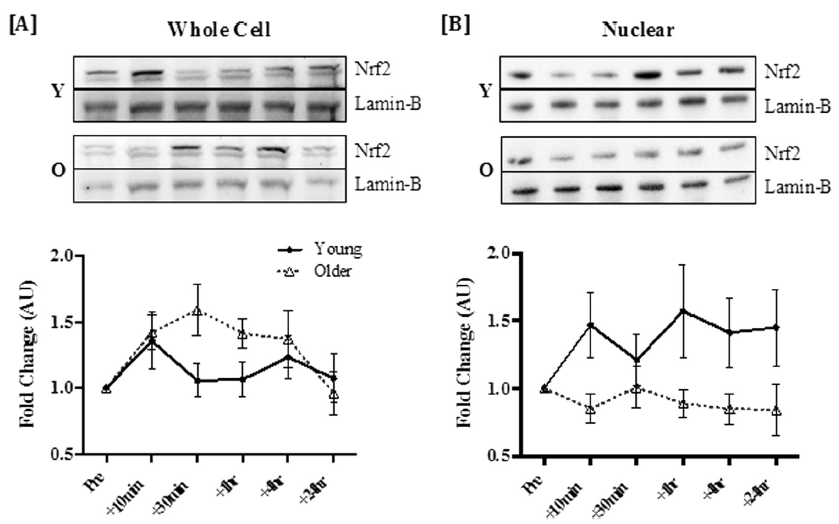
图2 运动在全细胞水平上诱导Nrf2增加,但在老年人中细胞核积累减弱。运动后PBMCs全细胞[A]和细胞核部分[B]中Nrf2的响应。数值为平均值±SEM。在相应图的上方提供了轻人(Y)和老年人(O)的Nrf2变化的代表性条带和内参条带。全细胞Nrf2的倍数变化主要受时间影响(P=0.003)。细胞核聚积只在年轻人中显著增加,在老年人中则不显著(主要影响因素年龄,p=0.031)。经Elsevier许可转载。
另外两项人类研究虽然没有直接研究Nrf2信号传导,但与之相关,两项研究包括了测量响应急性运动的Nrf2的mRNA。在含氧量正常的恢复条件下,经过90分钟的急性自行车运动后,年轻健康男性骨骼肌中Nrf2和靶向SOD2的基因表达显著增加。有趣的是,在缺氧的恢复条件下未发现这些反应。同样地,在跑步机中等强度运动2小时30分钟后,经常运动的中年妇女Nrf2 的mRNA显著增加;然而,久坐妇女在运动前后Nrf2或靶点的基因表达没有变化,这表明随着年龄的增长,健身可能在维持急性Nrf2响应中起到作用。
与细胞培养研究类似,一些动物研究表明,Nrf2信号的增加依赖于运动的持续时间。持续不到一小时的跑步机运动不会引起Nrf2的mRNA或蛋白质表达的变化。当持续时间增加到90分钟或更长时,骨骼肌组织和脑匀浆中的Nrf2激活变得明显,骨骼肌中Nrf2蛋白表达显著增加,两种组织中的基因表达显著增加。在耐力运动的一个极端例子中,Li和同事证明,在跑步机上跑1小时小鼠的Nrf2活性没有差异,但跑6小时后,Nrf2细胞核聚积和Nrf2-ARE的结合强劲增加。且与非运动对照组和1小时组相比,多种II相酶的基因表达显著增加。与这些结果相反,其他研究报道在跑步机运动1小时和中等强度循环30分钟后,Nrf2蛋白质丰度增加。这些研究之间观察到的差异可能是方案强度差异的结果(总结在表1中)。
目前还没有研究运动强度对Nrf2信号响应的影响。然而,Wang等人发现运动诱导的线粒体H2O2含量与Nrf2基因和蛋白表达之间存在正相关(根据运动不同时间的反应进行测量)。然而,假设高强度导致更大的氧化应激;这些数据以及肌管中电脉冲刺激的结果表明强度可能影响Nrf2信号。运动的刺激效应(无论是持续时间还是强度)可能有一个上限,因为运动到筋疲力尽的大鼠没有表现出Nrf2蛋白或II相酶蛋白水平或包括谷胱甘肽还原酶(GR)、NQO1和谷胱甘肽-s-转移酶(GST)在内的酶活性增加,除非它们之前接受过萝卜硫素,一种有效的Nrf2活化剂。
另一个需要考虑的机制来自于Xue等人最近的研究,其证明在刺激II相抗氧化物表达方面,Nrf2的积累可能不如Nrf2进出细胞核的频率重要。在他们的模型中,Nrf2循环的速率和振幅决定了下游信号的强度——尽管很难解读这些体外数据,但这个概念很有趣。运动的方式、强度和持续时间都可能影响体内Nrf2循环的速率和振幅。例如,即使最短持续时间为30秒的最大运动也会引起氧化还原平衡的系统性变化;随着强度和持续时间的增加,氧化还原平衡的变化幅度也会增加。因此,与传统的有氧运动相比,通过高强度间歇训练进行重复的氧化还原平衡移位可能会对Nrf2的进出频率产生不同的影响。确定Nrf2激活的最佳“剂量”或运动量为未来研究提供了巨大前景。
4、Nrf2信号对定期运动的响应
与急性运动相比,定期运动训练对Nrf2响应的影响已被广泛研究。表2概述了这些研究。无论持续时间(4-24周)或训练方案(传统的中等强度或高强度间歇训练)如何,啮齿类动物模型中的定期有氧运动始终被证明能够激活包括骨骼肌、肾脏、大脑、肝脏、睾丸、前列腺和心肌在内的多种组织的Nrf2信号。
表2 定期运动诱导Nrf2信号
表2 (续表)

NR未报告或未测量,SMT骨骼肌组织,PBMC外周血单核细胞,NUC核,WC全细胞。
综上所述,这些动物研究的数据表明,定期运动会上调Nrf2蛋白丰度和II相酶的数量和/或酶的活性。迄今为止,还没有对人体内Nrf2信号进行运动干预的研究。然而,一项比较久坐和活动老年人的单次肌肉活检中Nrf2和Keap1蛋白含量的横断面研究表明,活动老年人的Nrf2蛋白含量明显更高,Nrf2/Keap1比率更高,表明定期运动可能会减弱Nrf2信号的年龄相关变化。运动干预是否能恢复Nrf2信号传导已受损个体的氧化还原平衡仍是个问题。来自老年动物的运动干预数据支持老年人有恢复Nrf2信号的潜力。另一个有趣的问题是,一个人在一生中开始运动的时间是否重要。最近的两项研究,虽然没有测量Nrf2信号,但证明老年人的终身训练与骨骼肌中氧化还原平衡的年龄相关变化的代偿适应有关。Zhao等人在加速衰老的小鼠模型中(SAMP8)更直接地研究了这个问题。他们有3组动物接受游泳运动,分别是终身的、在生命周期早期开始的和晚期开始的。这些组还与非运动对照组进行了比较。总的来说,晚期组仍然比久坐组有一些好处,这支持了一句古老的格言,即开始运动永远不会太迟。然而,与晚期组相比,终身组和早期组显示出明显更大的益处,这表明早期开始运动在运动诱导信号的年龄相关缺陷方面提供了更大保护。重要的是,有证据表明经常运动可以增强对之后损害的恢复力。例如,接受过跑步机训练或保持静坐状态的大鼠,脑内输注药物破坏多巴胺受体以模拟帕金森氏病。运动训练可保护大鼠免受神经退行性病变、氧化应激和药物反应性Nrf2降低的影响。此外,当Nrf2被阻断时,运动保护作用并不明显,这支持了Nrf2在运动介导的神经保护中的作用。
据我们所知,目前为止还没有任何研究针对传统抗阻力训练来研究Nrf2信号。 Gomes等人采用带负重背心的跳跃方案作为对Sprague-Dawley大鼠进行抗阻力训练(也称阻力训练,译者注)。在幼年动物中没有观察到Nrf2表达的变化,而接受相同训练方案的老年动物Nrf2表达降低。老年动物的Nrf2表达基线水平明显高于年轻动物,这部分解释了运动训练后Nrf2的下降。有可能是老年动物上调了Nrf2以抵消日益增长的促氧化环境。运动训练可以改善Nrf2诱导反应,从而降低基线值。可能需要测量运动干预前后对急性运动的反应,以阐明这些观察到的Nrf2表达差异。在接受过卧床休息的人身上也观察到了类似情况,不论他们是否接受过伴随的抗阻力训练。卧床休息控制(无运动)显示骨骼肌中Nrf2的mRNA水平升高;然而,卧床休息加上阻力训练与卧床前休息水平相比,没有引起Nrf2表达的变化,这表明阻力运动可维持氧化还原平衡。这两项研究都没有集中于Nrf2信号本身,只包括测量Nrf2基因表达变化。需要进一步的研究来阐明抗阻力训练是否能诱导Nrf2信号传导。
5、抗氧化剂补充对Nrf2信号传导的影响
虽然这些证据明确支持运动作为一种通过激活Nrf2诱导的内源性抗氧化防御的可行方法,但人们总是关注是否可以通过外源性补充剂或药物实现这一点。考虑到氧化应激在衰老中的作用,外源性抗氧化剂补充通过减缓衰老或活性氧产生速率,减少自由基与细胞成分接触,并促进更平衡的细胞氧化还原环境来,对抗由自由基引起的细胞损伤和与年龄相关的病理学变化。这一假设导致了大量的研究,研究了外源性补充剂与成功老化(快乐、满足、幸福地度过老化时期,在生理、心理和社会等方面都感到相对适应,译者摘注)和/或延长寿命之间的关系。事实上,对“抗氧化剂和衰老”关键词的公开搜索产生了超过13000份结果。尽管有大量的文献,但对于补充抗氧化剂的益处仍没有达成共识。许多研究表明补充抗氧化剂没有影响,一些研究甚至表明抗氧化剂补充剂的负面影响。目前,美国超过40%的成年人口每天至少摄入多维生素补充剂,仅在美国,每年就产生320亿美元的产业收入。研究中最常见的两种维生素是维生素C和E。
维生素C(抗坏血酸/抗坏血酸)是一种有效的亲水性抗氧化剂,能够直接中和活性氧,尽管速度非常慢。维生素E(α-生育酚)是细胞膜的主要疏水性抗氧化剂,是体内脂质过氧化的有力抑制剂。由于其疏水性,它被储存在细胞膜中,可以清除过氧化自由基,比过氧化自由基与细胞膜反应快得多。此外,α-生育酚能直接清除线粒体膜中的活性氧。鉴于维生素E的抗氧化特性,它被吹嘘具有通过抑制循环脂蛋白的氧化修饰进而预防动脉粥样硬化的潜力。然而,根据这些反应,维生素C和E可以提高细胞存活率,最近的荟萃分析表明,补充抗氧化维生素没有效果,甚至可能增加发病风险。运动和抗氧化维生素补充剂之间的相互作用使情况更加复杂。外源抗氧化剂可能通过在细胞信号传导开始前降低ROS来减弱信号传导(见图1)。
因此,活性氧的许多有益作用被中断,这与通过Nrf2介导的内源性抗氧化防御作用控制的活性氧下降形成了直接对比。在一项双盲、安慰剂对照的维生素C和E补充研究中,Morrison等人清楚地证明,通过测定F2异前列素对急性运动挑战的响应,补充剂对减少氧化应激没有好处。此外,他们还发现,在对使用补充剂的成人进行为期4周的运动干预后,SOD1和SOD2蛋白表达减弱,而安慰剂组这些蛋白的表达显著增加。同时补充剂的运动适应不良反应在功能水平上也表现出来,补充剂减少了疲劳试验、血压控制和葡萄糖耐量的改善。在肌肉水平上,补充抗氧化剂抑制了线粒体适应耐力训练的几个标志物;尽管还不清楚补充抗氧化剂是否会抑制人类的耐力训练表现。
6、运动和植物营养素Nrf2活化剂有协同作用吗?
深入讨论植物营养素或植物化学物质对Nrf2信号传导的影响超出了本综述的范围。相反,我们将关注最近的研究,这些研究已经证明了运动与已知的Nrf2活化剂结合在进一步改善氧化还原平衡方面的潜在协同效应。Pala等证明辅酶Q10(COQ10)补充剂和运动对雄性Wistar大鼠有协同作用。运动和辅酶Q10补充剂的作用相互独立,显著增加心脏、肝脏和骨骼肌中的Nrf2表达,超过久坐对照组。在所有组织中,补充剂和运动对Nrf2表达的联合作用都显著增强,并且类似模式在诱导HO-1中也观察到。Sprague-Dawley大鼠补充了已知的Nrf2激活剂——冬虫夏草,在15天的游泳计划中Nrf2蛋白表达和II相酶表达水平显著高于单独运动组的水平。这一领域值得进一步研究。图3显示了来自早期研究的数据,这些研究表明,运动效果是如何通过同时补充抗氧化剂而被抑制的,以及如何通过同时使用植物营养素Nrf2激活剂而得到增强的。
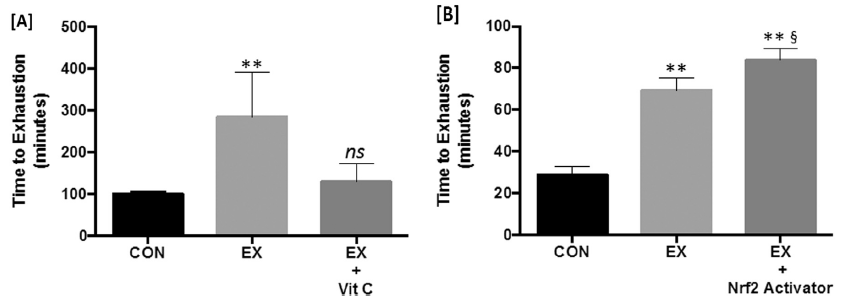
图3 运动结合补充抗氧化剂维生素的抑制运动适应,而Nrf2激活剂增强运动效果。与久坐对照组小鼠相比,接受维生素C补充并运动的啮齿动物未能改善其疲劳时间[A]。与之相反,与静坐对照组和非补充运动组相比,在疲劳时间试验的运动过程中,补充了Nrf2激活化合物的大鼠表现明显更好,表明其具有潜在的协同效应[B]。数据源自Gomez-Cabrera等人和Kumar等人。
激活Nrf2和II相反应的另一个可能机制是热量限制。据我们所知,目前还没有关于热量限制和运动结合对Nrf2和下游信号传导影响的随机对照研究。
7、结论与未来方向
总之,这些数据说明运动通过氧化还原激活Nrf2信号产生了许多好处。Nrf2在包括大脑、肾脏和睾丸在内各种组织中的激活可能是运动如何产生众所周知的,不仅限于骨骼肌和心肌的系统效应的一个重要机制。此外,还有一些新的数据表明,这些结果实际上可以转化到人类。显然,我们还需要更多的研究,特别是人体运动干预研究的随机对照试验。动物研究和人类研究几乎都局限于雄性/男性。结果需要扩展到雌性/女性,并解释可能的性别差异。在增强对Nrf2转录因子在健康和疾病中重要作用的理解方面,使用Nrf2基因敲除小鼠具有极其重要的价值。Nrf2−/−表现出与老龄小鼠非常相似的表型,表明Nrf2-Keap1信号的中断是生物老化的一个原因,这使得Nrf2成为一个有希望的靶点,特别是治疗生物老化以预防年龄相关疾病中。
动物研究的一个告诫是,通常只有两个时间点可用(急性运动前/后或运动干预前/后)。许多Nrf2调节基因和蛋白质可能有不同的时间-响应进程,因此可能需要多个时间点来捕捉整体变化。这些需要在未来进一步研究。
翻译:
Nrf2介导氧化还原作用对运动的适应
摘要综述的主要目的是总结当前关于急性运动和定期运动对核因子红细胞样2相关因子2(Nrf2)活性和Nrf2信号下游靶点的影响的文献。Nrf2(在人体内由NFE2L2基因编码)是抗氧化剂防御的主要调节因子,是一种调节200多个细胞保护基因表达的转录因子。越来越多的证据表明,Nrf2信号在氧化应激如何介导运动的有益作用中起着关键作用。通过急性运动诱导的氧化应激偶发性增加刺激了Nrf2活化,与定期运动一样重复刺激时,会导致内源性抗氧化防御能力与总体更大的抗氧化应激破坏性作用能力的上调。运动造成的不同组织Nrf2激活证据可能是运动如何发挥其众所周知的系统效应的一个重要机制,这些系统效应不仅限于骨骼肌和心肌。此外,还有一些动物研究成果转化为人类的数据。
1、简介
随着分子生物学的发展及其在运动中的应用,人们对于急性运动诱导细胞信号导致多次运动后积累生理适应性的认识得到了极大提高。急性运动激活初级信使,如一磷酸腺苷(AMP)、钙和机械应激,以及随后的次级信使,包括AMP活化蛋白激酶(AMPK)、钙/钙调蛋白依赖性蛋白激酶(CAMK)和p38有丝分裂原活化蛋白激酶,导致mRNA转录的急性变化。急性运动刺激的另一个重要的初级信使是氧化还原平衡(或氧化还原电位)的暂时性变化,通过产生活性氧物质(ROS)进入更加氧化的状态。虽然ROS最初被认为具有损伤性,但现在人们对ROS在细胞信号传导、免疫功能调节、基因转录和凋亡中的重要作用有了更深入的了解。一个主要的例子是ROS在转录因子核因子红细胞样2相关因子2(Nrf2)激活中的作用,其是抗氧化剂酶和细胞抗应激的主要调节因子。啮齿动物研究的数据表明,靶向Nrf2活性以降低疾病风险的治疗潜力。靶向Nrf2敲除(Nrf2–/–)模型在所有组织中对毒性和氧化应激源表现出抵抗力明显受损,因此Nrf2–/–小鼠更易患慢性疾病。靶向Nrf2激活的药理学和遗传学(负调节子敲除)方法有效地改善氧化应激源造成的损害并恢复对慢性疾病的抵抗。Nrf2信号中断的情况下病理学的增加,以及随后在Nrf2 活化剂存在下的逆转已经在其他地方进行了综述。
越来越多的证据表明,Nrf2途径在氧化应激如何调节运动的有益作用中起着关键作用。急性运动诱导的氧化应激的偶发性增加会刺激Nrf2激活,与定期运动一样重复刺激时,这可能会导致内源性抗氧化防御的上调,以及总体上更大的能力以抵消氧化对核酸、蛋白质和脂质的损伤作用。本综述的主要目的是总结当前关于急性运动和定期运动或运动干预对Nrf2活性和Nrf2信号下游靶点影响的文献。我们还研究了支持或反对与运动结合使用抗氧化补充剂和植物营养素Nrf2活化剂的证据。由于干预效果在目标受损时更容易观察到,我们将着重研究衰老作为受损细胞信号的非疾病模型,其通常与年龄相关的病理学相协调。我们从Nrf2调节的简要概述开始。
2、 Nrf2:细胞防御的主调节子
转录因子Nrf2(在人类中由NFE2L2基因编码)是抗氧化防御的主调节子,调节超过200种保护氧化应激的细胞保护基因。Nrf2是碱性亮氨酸拉链(bZIP)转录因子家族的一员,在无应激条件下通过与胞质溶胶中的Kelch样红系细胞相关CNC同源关联蛋白1(Keap1)的同二聚体结合而被抑制[17]。Nrf2和Keap1之间的相互作用在物种间高度保守,表明其调节作用的重要[18]。在这种状态下,Keap1作为Nrf2降解的衔接蛋白发挥作用,其由Cul3/Rbx1介导,促进Nrf2泛素化和随后26s蛋白酶体对Nrf2的降解。Keap1/Cul3/Rbx1也作为一种额外的负调节因子存在于细胞核中[19]。
半胱氨酸残基被以一种独特的形式修饰来响应氧化或亲电刺激,不同的结构诱导物与Keap1上不同的半胱氨酸残基组合反应,从而产生相同的生物响应,特别是从Nrf2脱离并激活Nrf2-抗氧化反应元件(ARE)响应,或通过稳定Keap1-Nrf2复合物,减缓蛋白酶体对其的降解[20–23]。一旦从Keap1上脱离,Nrf2就转移到细胞核中,在那里它与小Maf蛋白异二聚体化,并结合顺式作用元件AREs,有效地激活II相解毒酶的转录(见图1)。
图1 Nrf2信号通路。Nrf2被运动诱导的ROS或植物营养素Nrf2激动剂激活。抗氧化补充剂抑制了运动诱导的ROS的信号,从而抑制下游的Nrf2信号传导。
在治疗潜力方面,这种半胱氨酸编码作为多种化合物和刺激物的有效Nrf2诱导物,之间相互独立,引起了人们极大的兴趣。例如,运动诱导的Nrf2的活性氧活化可能通过氧化这些半胱氨酸残基发生。同样,某些植物营养素被证明能够激活Nrf2,这个过程可能是通过改变半胱氨酸残基而发生的,而这些残基不同于运动中的靶点。如果事实上是这样的话,运动训练和Nrf2激动化合物之间可能存在协同效应。一些最新数据支持这一点,这些数据在本综述的第6节中给出。对于那些对Nrf2激活机制的更详细背景感兴趣的人,我们请读者参考这方面近期的一篇优秀评论[16]。
值得注意的是,在某些情况下,如癌症,Nrf2的激活和信号传导可能会产生有害影响。在正常情况下,Nrf2信号可以防止癌症,正如十字花科蔬菜中的植物营养素刺激Nrf2活化的效果一样。然而,在癌细胞中,Nrf2的激活可能赋予细胞抗应激能力,使细胞不易受到化疗治疗的影响。研究表明,在癌症肿瘤中,Nrf2被上调,在化疗期间可能需要使用Nrf2抑制剂提高治疗效果。
3、Nrf2信号对急性运动的反应
使用C2C12骨骼肌细胞进行的细胞培养为证明Nrf2被ROS激活的研究提供了证据,当抗氧化剂(如N-乙酰半胱氨酸)应用于培养基时,激活被抑制。Horie等人使用电脉冲刺激(EPS)模拟急性运动,并证明Nrf2表达与刺激强度和持续时间相关。此外,EPS刺激了Nrf2相关抗氧化基因的表达,当siRNA转染敲低细胞中的Nrf2时,表达减弱。其他研究表明,用H2O2处理肌管或大鼠心肌细胞后,Nrf2蛋白表达增加。
这些体外结果转化为体内结果主要在动物模型中完成,最近的一项研究是在临床研究。表1列出了这些研究。值得注意的是,早期的一些研究在其免疫印迹中针对一个非特异性的Nrf2(57 kDa)条带。尽管预测的Nrf2分子量为57 kDa,但一些研究小组已经确定90-110 kDa带是与生物相关的Nrf2靶点。
表1 急性运动诱导Nrf2信号
NR未报告或未测量,SMT骨骼肌组织,PBMC外周血单核细胞,NUC核,WC全细胞。
在野生型小鼠中,单次急性运动提高了骨骼肌中的Nrf2基因表达和其蛋白丰度,以及依赖Nrf2的II相酶。在另外两项测量运动对小鼠心肌中Nrf2信号传导影响的研究中,结果基本上与骨骼肌研究的数据一致。然而,这些研究使用了“急性运动应激”,其为连续两天60分钟的跑步机运动。虽然很明显时间不足以归类为运动训练,但众所周知,连续运动会导致mRNA表达的累积反应,可能需要观察蛋白质丰度的可测量变化[44]。然而,Muthusamy等人[42]提供了对运动反应的Nrf2信号的最全面分析,并比较了年轻野生型(WT)和Nrf2–/–小鼠。尽管两组的ROS都有相似的增加,但运动引起的核积累显著增加,并且Nrf2在WT小鼠心肌中结合,而在Nrf2–/–小鼠中没有观察到Nrf2活性的变化。在WT小鼠中几乎所有II相酶的基因表达都显著增加,唯一的例外是血红素加氧酶-1(HO-1)。II相抗氧化蛋白的表达变化不太一致,仅在HO-1、谷氨酸半胱氨酸连接酶(GCLM)和葡萄糖-6-磷酸脱氢酶(G6PD)中观察到变化。
关于运动诱导的人体Nrf2信号传导的数据仍非常有限,但最近的一项研究首次表明,在年轻和老年人的外周血单核细胞(PBMCs)中,急性运动增加了全细胞水平的Nrf2蛋白丰度[45]。此外,仅在年轻组中观察到Nrf2的核聚积,表明老化与Nrf2的核输入受损有关(见图2)。毫不奇怪,运动刺激了年轻人而不是老年人的HO-1和NAD(P)H醌脱氢酶1(NQO1)基因表达的增加。之前在小鼠心肌和大鼠肝脏中显示了与年龄相关的胞质溶胶和细胞核中Nrf2水平以及Nrf2/ARE结合能力的降低,并伴随响应急性运动的Nrf2依赖的抗氧化基因表达的减弱。
图2 运动在全细胞水平上诱导Nrf2增加,但在老年人中细胞核积累减弱。运动后PBMCs全细胞[A]和细胞核部分[B]中Nrf2的响应。数值为平均值±SEM。在相应图的上方提供了轻人(Y)和老年人(O)的Nrf2变化的代表性条带和内参条带。全细胞Nrf2的倍数变化主要受时间影响(P=0.003)。细胞核聚积只在年轻人中显著增加,在老年人中则不显著(主要影响因素年龄,p=0.031)。经Elsevier许可转载。
另外两项人类研究虽然没有直接研究Nrf2信号传导,但与之相关,两项研究包括了测量响应急性运动的Nrf2的mRNA。在含氧量正常的恢复条件下,经过90分钟的急性自行车运动后,年轻健康男性骨骼肌中Nrf2和靶向SOD2的基因表达显著增加。有趣的是,在缺氧的恢复条件下未发现这些反应。同样地,在跑步机中等强度运动2小时30分钟后,经常运动的中年妇女Nrf2 的mRNA显著增加;然而,久坐妇女在运动前后Nrf2或靶点的基因表达没有变化,这表明随着年龄的增长,健身可能在维持急性Nrf2响应中起到作用。
与细胞培养研究类似,一些动物研究表明,Nrf2信号的增加依赖于运动的持续时间。持续不到一小时的跑步机运动不会引起Nrf2的mRNA或蛋白质表达的变化。当持续时间增加到90分钟或更长时,骨骼肌组织和脑匀浆中的Nrf2激活变得明显,骨骼肌中Nrf2蛋白表达显著增加,两种组织中的基因表达显著增加。在耐力运动的一个极端例子中,Li和同事证明,在跑步机上跑1小时小鼠的Nrf2活性没有差异,但跑6小时后,Nrf2细胞核聚积和Nrf2-ARE的结合强劲增加。且与非运动对照组和1小时组相比,多种II相酶的基因表达显著增加。与这些结果相反,其他研究报道在跑步机运动1小时和中等强度循环30分钟后,Nrf2蛋白质丰度增加。这些研究之间观察到的差异可能是方案强度差异的结果(总结在表1中)。
目前还没有研究运动强度对Nrf2信号响应的影响。然而,Wang等人发现运动诱导的线粒体H2O2含量与Nrf2基因和蛋白表达之间存在正相关(根据运动不同时间的反应进行测量)。然而,假设高强度导致更大的氧化应激;这些数据以及肌管中电脉冲刺激的结果表明强度可能影响Nrf2信号。运动的刺激效应(无论是持续时间还是强度)可能有一个上限,因为运动到筋疲力尽的大鼠没有表现出Nrf2蛋白或II相酶蛋白水平或包括谷胱甘肽还原酶(GR)、NQO1和谷胱甘肽-s-转移酶(GST)在内的酶活性增加,除非它们之前接受过萝卜硫素,一种有效的Nrf2活化剂。
另一个需要考虑的机制来自于Xue等人最近的研究,其证明在刺激II相抗氧化物表达方面,Nrf2的积累可能不如Nrf2进出细胞核的频率重要。在他们的模型中,Nrf2循环的速率和振幅决定了下游信号的强度——尽管很难解读这些体外数据,但这个概念很有趣。运动的方式、强度和持续时间都可能影响体内Nrf2循环的速率和振幅。例如,即使最短持续时间为30秒的最大运动也会引起氧化还原平衡的系统性变化;随着强度和持续时间的增加,氧化还原平衡的变化幅度也会增加。因此,与传统的有氧运动相比,通过高强度间歇训练进行重复的氧化还原平衡移位可能会对Nrf2的进出频率产生不同的影响。确定Nrf2激活的最佳“剂量”或运动量为未来研究提供了巨大前景。
4、Nrf2信号对定期运动的响应
与急性运动相比,定期运动训练对Nrf2响应的影响已被广泛研究。表2概述了这些研究。无论持续时间(4-24周)或训练方案(传统的中等强度或高强度间歇训练)如何,啮齿类动物模型中的定期有氧运动始终被证明能够激活包括骨骼肌、肾脏、大脑、肝脏、睾丸、前列腺和心肌在内的多种组织的Nrf2信号。
表2 定期运动诱导Nrf2信号
表2 (续表)
NR未报告或未测量,SMT骨骼肌组织,PBMC外周血单核细胞,NUC核,WC全细胞。
综上所述,这些动物研究的数据表明,定期运动会上调Nrf2蛋白丰度和II相酶的数量和/或酶的活性。迄今为止,还没有对人体内Nrf2信号进行运动干预的研究。然而,一项比较久坐和活动老年人的单次肌肉活检中Nrf2和Keap1蛋白含量的横断面研究表明,活动老年人的Nrf2蛋白含量明显更高,Nrf2/Keap1比率更高,表明定期运动可能会减弱Nrf2信号的年龄相关变化。运动干预是否能恢复Nrf2信号传导已受损个体的氧化还原平衡仍是个问题。来自老年动物的运动干预数据支持老年人有恢复Nrf2信号的潜力。另一个有趣的问题是,一个人在一生中开始运动的时间是否重要。最近的两项研究,虽然没有测量Nrf2信号,但证明老年人的终身训练与骨骼肌中氧化还原平衡的年龄相关变化的代偿适应有关。Zhao等人在加速衰老的小鼠模型中(SAMP8)更直接地研究了这个问题。他们有3组动物接受游泳运动,分别是终身的、在生命周期早期开始的和晚期开始的。这些组还与非运动对照组进行了比较。总的来说,晚期组仍然比久坐组有一些好处,这支持了一句古老的格言,即开始运动永远不会太迟。然而,与晚期组相比,终身组和早期组显示出明显更大的益处,这表明早期开始运动在运动诱导信号的年龄相关缺陷方面提供了更大保护。重要的是,有证据表明经常运动可以增强对之后损害的恢复力。例如,接受过跑步机训练或保持静坐状态的大鼠,脑内输注药物破坏多巴胺受体以模拟帕金森氏病。运动训练可保护大鼠免受神经退行性病变、氧化应激和药物反应性Nrf2降低的影响。此外,当Nrf2被阻断时,运动保护作用并不明显,这支持了Nrf2在运动介导的神经保护中的作用。
据我们所知,目前为止还没有任何研究针对传统抗阻力训练来研究Nrf2信号。 Gomes等人采用带负重背心的跳跃方案作为对Sprague-Dawley大鼠进行抗阻力训练(也称阻力训练,译者注)。在幼年动物中没有观察到Nrf2表达的变化,而接受相同训练方案的老年动物Nrf2表达降低。老年动物的Nrf2表达基线水平明显高于年轻动物,这部分解释了运动训练后Nrf2的下降。有可能是老年动物上调了Nrf2以抵消日益增长的促氧化环境。运动训练可以改善Nrf2诱导反应,从而降低基线值。可能需要测量运动干预前后对急性运动的反应,以阐明这些观察到的Nrf2表达差异。在接受过卧床休息的人身上也观察到了类似情况,不论他们是否接受过伴随的抗阻力训练。卧床休息控制(无运动)显示骨骼肌中Nrf2的mRNA水平升高;然而,卧床休息加上阻力训练与卧床前休息水平相比,没有引起Nrf2表达的变化,这表明阻力运动可维持氧化还原平衡。这两项研究都没有集中于Nrf2信号本身,只包括测量Nrf2基因表达变化。需要进一步的研究来阐明抗阻力训练是否能诱导Nrf2信号传导。
5、抗氧化剂补充对Nrf2信号传导的影响
虽然这些证据明确支持运动作为一种通过激活Nrf2诱导的内源性抗氧化防御的可行方法,但人们总是关注是否可以通过外源性补充剂或药物实现这一点。考虑到氧化应激在衰老中的作用,外源性抗氧化剂补充通过减缓衰老或活性氧产生速率,减少自由基与细胞成分接触,并促进更平衡的细胞氧化还原环境来,对抗由自由基引起的细胞损伤和与年龄相关的病理学变化。这一假设导致了大量的研究,研究了外源性补充剂与成功老化(快乐、满足、幸福地度过老化时期,在生理、心理和社会等方面都感到相对适应,译者摘注)和/或延长寿命之间的关系。事实上,对“抗氧化剂和衰老”关键词的公开搜索产生了超过13000份结果。尽管有大量的文献,但对于补充抗氧化剂的益处仍没有达成共识。许多研究表明补充抗氧化剂没有影响,一些研究甚至表明抗氧化剂补充剂的负面影响。目前,美国超过40%的成年人口每天至少摄入多维生素补充剂,仅在美国,每年就产生320亿美元的产业收入。研究中最常见的两种维生素是维生素C和E。
维生素C(抗坏血酸/抗坏血酸)是一种有效的亲水性抗氧化剂,能够直接中和活性氧,尽管速度非常慢。维生素E(α-生育酚)是细胞膜的主要疏水性抗氧化剂,是体内脂质过氧化的有力抑制剂。由于其疏水性,它被储存在细胞膜中,可以清除过氧化自由基,比过氧化自由基与细胞膜反应快得多。此外,α-生育酚能直接清除线粒体膜中的活性氧。鉴于维生素E的抗氧化特性,它被吹嘘具有通过抑制循环脂蛋白的氧化修饰进而预防动脉粥样硬化的潜力。然而,根据这些反应,维生素C和E可以提高细胞存活率,最近的荟萃分析表明,补充抗氧化维生素没有效果,甚至可能增加发病风险。运动和抗氧化维生素补充剂之间的相互作用使情况更加复杂。外源抗氧化剂可能通过在细胞信号传导开始前降低ROS来减弱信号传导(见图1)。
因此,活性氧的许多有益作用被中断,这与通过Nrf2介导的内源性抗氧化防御作用控制的活性氧下降形成了直接对比。在一项双盲、安慰剂对照的维生素C和E补充研究中,Morrison等人清楚地证明,通过测定F2异前列素对急性运动挑战的响应,补充剂对减少氧化应激没有好处。此外,他们还发现,在对使用补充剂的成人进行为期4周的运动干预后,SOD1和SOD2蛋白表达减弱,而安慰剂组这些蛋白的表达显著增加。同时补充剂的运动适应不良反应在功能水平上也表现出来,补充剂减少了疲劳试验、血压控制和葡萄糖耐量的改善。在肌肉水平上,补充抗氧化剂抑制了线粒体适应耐力训练的几个标志物;尽管还不清楚补充抗氧化剂是否会抑制人类的耐力训练表现。
6、运动和植物营养素Nrf2活化剂有协同作用吗?
深入讨论植物营养素或植物化学物质对Nrf2信号传导的影响超出了本综述的范围。相反,我们将关注最近的研究,这些研究已经证明了运动与已知的Nrf2活化剂结合在进一步改善氧化还原平衡方面的潜在协同效应。Pala等证明辅酶Q10(COQ10)补充剂和运动对雄性Wistar大鼠有协同作用。运动和辅酶Q10补充剂的作用相互独立,显著增加心脏、肝脏和骨骼肌中的Nrf2表达,超过久坐对照组。在所有组织中,补充剂和运动对Nrf2表达的联合作用都显著增强,并且类似模式在诱导HO-1中也观察到。Sprague-Dawley大鼠补充了已知的Nrf2激活剂——冬虫夏草,在15天的游泳计划中Nrf2蛋白表达和II相酶表达水平显著高于单独运动组的水平。这一领域值得进一步研究。图3显示了来自早期研究的数据,这些研究表明,运动效果是如何通过同时补充抗氧化剂而被抑制的,以及如何通过同时使用植物营养素Nrf2激活剂而得到增强的。
图3 运动结合补充抗氧化剂维生素的抑制运动适应,而Nrf2激活剂增强运动效果。与久坐对照组小鼠相比,接受维生素C补充并运动的啮齿动物未能改善其疲劳时间[A]。与之相反,与静坐对照组和非补充运动组相比,在疲劳时间试验的运动过程中,补充了Nrf2激活化合物的大鼠表现明显更好,表明其具有潜在的协同效应[B]。数据源自Gomez-Cabrera等人和Kumar等人。
激活Nrf2和II相反应的另一个可能机制是热量限制。据我们所知,目前还没有关于热量限制和运动结合对Nrf2和下游信号传导影响的随机对照研究。
7、结论与未来方向
总之,这些数据说明运动通过氧化还原激活Nrf2信号产生了许多好处。Nrf2在包括大脑、肾脏和睾丸在内各种组织中的激活可能是运动如何产生众所周知的,不仅限于骨骼肌和心肌的系统效应的一个重要机制。此外,还有一些新的数据表明,这些结果实际上可以转化到人类。显然,我们还需要更多的研究,特别是人体运动干预研究的随机对照试验。动物研究和人类研究几乎都局限于雄性/男性。结果需要扩展到雌性/女性,并解释可能的性别差异。在增强对Nrf2转录因子在健康和疾病中重要作用的理解方面,使用Nrf2基因敲除小鼠具有极其重要的价值。Nrf2−/−表现出与老龄小鼠非常相似的表型,表明Nrf2-Keap1信号的中断是生物老化的一个原因,这使得Nrf2成为一个有希望的靶点,特别是治疗生物老化以预防年龄相关疾病中。
动物研究的一个告诫是,通常只有两个时间点可用(急性运动前/后或运动干预前/后)。许多Nrf2调节基因和蛋白质可能有不同的时间-响应进程,因此可能需要多个时间点来捕捉整体变化。这些需要在未来进一步研究。
本文由福山生物整理翻译,转载请注明出处。