






最新资讯



-
电话: 400-113-6988 邮箱: dongfangxicao@163.com 地址: 深圳市南山区粤海街道高新中三道9号环球数码大厦19楼
↓ 微信扫码识别关注我们 ↓ 
miR-483-3P上调DKK3在黑树莓花色苷介导的结直肠癌化学预防中的重要作用
发表于:2021-02-20 作者:admin 来源:本站 点击量:26622
原文:Guo J, Yang Z, Zhou H, et al. Upregulation of DKK3 by miR‐483‐3p plays an important role in the chemoprevention of colorectal cancer mediated by black raspberry anthocyanins. Molecular Carcinogenesis. 2019;1–11. https://doi.org/10.1002/mc.23138
翻译:
据报道,黑树莓(BRB)花色苷可作为一种潜在的结直肠癌(CRC)化学预防药物。然而,BRB花色苷抑制大肠癌细胞癌变的潜在机制尚未阐明。靶向重要抑癌基因的microRNAs(MiRNAs)的异常表达通常与大肠癌的发生有关。在本研究中,我们探讨了BRB花色苷在偶氮甲烷(AOM)/葡聚糖硫酸钠(DSS)诱导的小鼠大肠癌模型和人大肠癌细胞系中是否影响某些miRNAs的表达。采用miRNA微阵列分析方法,研究饲喂不含或不含BRB花青素的饲料诱导的AOM/DSS小鼠miRNA表达的差异。一种特殊的miRNA,miR-483-3p的表达在AOM/DSS诱导的小鼠中被发现在饲料中添加BRB花青素后显著降低。随后的实时定量聚合酶链反应和Western blot分析表明,miR-483-3p表达降低的同时,生物信息学分析预测的miR-483-3p的潜在靶点Dickkopf3(DKK3)的表达增加。当miR-483-3p特异性抑制剂下调miR-483-3p的表达水平时,DKK3的蛋白和信使RNA水平明显上调,提示DKK3可能是miR-483-3p的靶基因。此外,包括Wnt/β-catenin在内的DKK3信号通路下游因子也在MIR483-3p介导的BRB花色苷抗癌作用中发挥作用。因此,miR-483-3p可能是BRB花色苷介导的预防大肠癌的潜在靶点。
关键词:
BRB花色苷、结直肠癌、DKK3、miR-483-3p、Wnt/β-catenin通路
1、简介
MicroRNAs(MiRNAs)是一种小分子单链RNA,普遍存在于动植物中。它通过与靶基因mRNA的3’-非翻译区(3’-UTR)结合,抑制翻译或直接降解信使RNA(MRNA),直接影响细胞生长、器官形成、能量代谢、细胞增殖、凋亡和防御系统。miRNAs还参与癌症的发展,既可以是癌基因,也可以是抑癌基因。例如,miR-21-5p和miR-142-5p既可以作为致癌基因也可以作为抑癌基因,而miR-22、miR-490-3p和miR-93都可以作为肿瘤抑制基因。此外,miRNAs还可以作为检测癌症的有用标记物。例如,血液中的miR-193a-3p、miR-23a和miR-338-5p可用于大肠癌(CRC)的早期检测。miR-483-3p通过抑制DPC4/Smad4的表达发挥促癌作用,从而促进胰腺癌的细胞增殖和克隆性。随着越来越多的miRNAs被发现在结直肠癌中异常表达,miRNA的应用将成为结直肠癌诊断、靶向治疗和预后研究的热点。大肠癌是一种常见的癌症,是世界上第四大常见死亡原因。大肠癌的发生发展涉及多个基因和信号通路的改变。大肠癌的发生发展伴随着Wnt/β-catenin信号通路的异常激活。Dickkopf蛋白家族由DKK1、DKK2、DKK3和DKK4组成,是Wnt/β-catenin信号通路的细胞外抑制因子,它通过与LRP5/LRP5受体相互作用来影响Wnt信号通路的非经典激活。
长期食用黑树莓(BRB)可以降低患癌症、心脏病、关节炎和呼吸系统疾病的风险。食用BRB会产生更多的次生代谢物,从而改善健康。BRB花色苷是BRB在体内产生的主要代谢物之一,也是膳食中花色苷的主要来源。BRB最显著的有益作用是其潜在的癌症化学预防作用,已有多项研究对此进行了研究。我们先前的研究发现,BRB花色苷可以调节肠道微生物区系,进一步使分泌的卷曲相关蛋白5的启动子去甲基化,最终延缓偶氮甲烷/葡聚糖硫酸钠(AOM/DSS)处理小鼠的结直肠癌的发展。花色苷似乎是临床试验中使用的冷冻干燥BRB粉末中的关键抗癌成分。
我们最近的研究发现miR-24-1-5p可以下调Wnt/β-catenin信号通路,有助于其在预防大肠癌的发生发展中发挥作用,从而参与BRB花色苷介导的结直肠癌的化学预防过程中的miRNAs的作用。在本研究中,miR-24-1-5p可下调Wnt/miRNAs-catenin信号通路的表达,从而起到预防大肠癌发生的作用。在这项研究中,我们研究了另一种miRNA miR-483-3p的作用,以确定它可能通过口服BRB花色苷抑制AOM/DSS诱导的CRC小鼠模型中CRC的发展的机制。在用BRB花色苷处理人大肠癌细胞后,还检测了miR-483-3p的作用。
2、材料和方法
2.1收购BRB花青素
BRBs含有各种化学物质,如酚酸和花色苷。BRB花色苷的主要成分为花青素-3-O-葡萄糖苷、花青素-3-O-芦丁苷、花青素-3-O-木糖基-芦丁苷等。本研究采用JF Natural公司(天津建峰天然产品研发有限公司)提取BRB花色苷。简单地说,冷冻干燥的BRB粉末用乙醇-水(75:25,体积/体积)在35°C下搅拌3h,直至粉末变白,得到粗提物。萃取物经滤纸过滤后装入聚酰胺色谱柱分离成不同组分。进一步用制备高效液相色谱分离纯化得到纯化的BRB花色苷。BRB花色苷保存在−20°C的环境中。
2.2 CRC小鼠模型的建立
将20只C57雄性小鼠(体重18~20g,辽宁长生实验动物生物技术有限公司)随机分为2组,每组10只。其中一组被指定为模型组,按正常饮食喂养。另一组为试验组,饲喂含7.0molmol/g BRB花色苷的普通饲料。饲料中BRB花色苷的浓度是在我们前期研究的基础上选择的。两组小鼠在实验的第一天注射AOM(10 mg/kg),然后在实验的第8天再注射。同时,第1周每天用2%DSS水治疗,随后2周每天用正常水治疗,整个过程重复3次。因此,整个实验持续了9周,在治疗期结束时,处死并取出它们的结肠,并将其冷冻在液氮中。部分组织用于miRNA阵列分析,其余组织保存在−80°C进行进一步分析。
在整个实验过程中,小鼠被关在笼子里(每笼3~5只),保持在22°C±1°C和50%±5%的湿度下,光照-黑暗周期为12小时(上午7:00-晚上7:00)。所有的程序都是根据国家卫生研究院关于动物护理和使用的规定进行的,并得到了辽宁中医药大学伦理委员会的批准。
2.3 miRNA微阵列
提取的结肠组织进行微阵列分析。使用安捷伦(安捷伦,Palo Alto,CA)对原始数据中不同基因的表达进行微阵列(8*60K,设计ID:070155)和基本分析。
2.4 细胞培养
人HCT116和HT29大肠癌细胞系购自中国科学院(上海)细胞库。LOVO细胞和SW480细胞购自上海明景生物研究所。细胞培养在37°C的5%CO2加湿环境中。
2.5 miRNAs瞬时转染
miR-483-3P抑制剂和miR-NC购自GenePharma(中国上海)。LoVo和SW480细胞的瞬时转染按照制造商方案进行。
2.6 RNA提取及qRT-PCR
用TRIzol试剂(上海桑贡生物科技有限公司)从细胞和组织中提取总RNA。编码miRNA的第一链互补DNA(cDNA)用miRNA cDNA合成试剂盒(ComWin Biotech Co,Ltd,China,China)反转录获得。用ComWin Biotech Co,Ltd的miRNA实时检测试剂盒进行miR-483-3p的实时定量聚合酶链反应(qRT-PCR)。用PrimeScript RT试剂盒检测DKK3的mRNA水平,试剂盒采用gDNA擦除器和TB Green PreMix Ex Taq II(北京塔卡拉生物医学技术有限公司)。用特异引物(表S1)进行qRT-PCR,并使用应用生物系统7500实时PCR系统进行扩增。
2.7 蛋白质提取及Western blot分析
冰冻条件下先用放射免疫沉淀分析缓冲液(北京鼎国长生生物科技有限公司)裂解细胞,随后细胞裂解液离心。然后,使用针对DKK3、β-肌动蛋白、Bcl-2、BAX(Sangon Biotech Shanghai Co,Ltd)、Cyclin D1、c-Myc、CDK4或E-cadherin(细胞信号技术,MA)的抗体,对所获得的含有可溶性蛋白的上清液进行Western blot分析。使用Image J软件对条带强度进行量化。
2.8 伤口愈合试验
这些细胞在一个六孔培养板中培养,直到它们达到60%的融合。然后用无菌微吸管刮除细胞层形成伤口,并用新鲜培养基更换培养液。然后,将miRNAs转染细胞24小时,拍摄细胞层的显微照片。光镜下计算细胞迁移程度。
2.9 3-(4,5-二甲基噻唑-2-基)-2,5-二苯基四唑溴化法
BRB花色苷的原液用培养基稀释到合适的浓度,然后加入96孔板,其中包含生长到60%融合的细胞。在细胞中分别加入BRB花色苷到最终浓度为25和50μg/mL。对照组仅向细胞中加入等量培养基,而不加入BRB花色苷。用3-(4,5-二甲基噻唑-2-基)-2,5-二苯基四唑溴化物(Sangon Biotech Shanghai Co,Ltd)试剂(Sangon Biotech Shanghai Co,Ltd)测定细胞增殖。
2.10 miRNA靶基因的鉴定及其与大肠癌患者生存率的关系
利用在线数据库microRNA.org预测miR-483-3p的靶基因(http://www.microrna.org/microrna/home.do)。从人类蛋白质图谱网站(www.proteinatlas.org)获得结直肠癌患者结肠组织中DKK3的表达谱。并用miRbase(http://www.mirbase.org/)对不同物种的miR-483-3p序列进行比对。使用在线数据库PROGeneV2(http://genomics.jefferson.edu/proggene/)验证DKK3的表达与大肠癌患者生存率的关系。
2.11 统计方式
采用SPSS 19.0统计软件进行数据分析。不同组间比较采用t检验和方差分析。在P<0.05和P<0.01水平上有统计学意义。
3 结果
3.1 miR-483-3p可能在BRB花色苷介导的大肠癌预防中发挥作用
应用miRNA微阵列分析方法获得了AOM/DSS诱导的小鼠结肠中miRNA的表达特征,这些小鼠分别饲喂了不添加(对照)或添加BRB花色苷的饲料。在接受BRB花色苷组中,几个miRNAs的表达模式与对照组不同。接受BRB花色苷的小鼠显示其结肠中miR-483-3p的表达减少(图1A,B)。此外,qRT-PCR显示,经BRB花色苷处理的CRC细胞与未处理的CRC细胞相比,miR-483-3p的表达也降低(图1C和1F)。提示miR-483-3p可能在BRB花色苷介导的化学预防大肠癌中起作用。
3.2 DKK3是miR-483-3p的预测靶基因,DKK3的表达水平与大肠癌患者有关
为了更好地了解miR-483-3p在大肠癌中的意义,我们对不同物种的miR-483序列进行了比对。结果显示,高等哺乳动物的miR-483序列具有高度的保守性(图2A)。此外,生物信息学分析预测DKK3可能是miR-483-3p的潜在靶基因(图2B)。然后利用在线数据库PROGeneV2分析DKK3与大肠癌患者生存时间的关系。DKK3 mRNA水平与患者总生存率呈正相关(图2C)。此外,从人类蛋白图谱数据库网站获得的DKK3蛋白免疫组织化学染色数据显示,这些大肠癌患者中DKK3的表达下调(图2D,E)。综上所述,DKK3可能是miR-483-3p在大肠癌患者中低表达的靶基因。BRB花色苷可能通过下调miR-483-3p的表达来调节DKK3的表达。
为了进一步检测miR-483-3p对DKK3的作用,用miR-483-3p抑制剂转染LoVo细胞和SW480细胞,该抑制剂由与miR-483-3p序列互补的寡核苷酸组成。转染miR-483-3p抑制剂后,细胞中DKK3的蛋白和mRNA水平均显著升高(图3A-C和3F)。此外,经BRB花色苷处理的LoVo和SW480细胞显示DKK3蛋白水平升高(图3g,H),进一步支持DKK3可能是miR-483-3p的靶基因的推测,BRB花色苷可能通过下调miR-483-3p表达来下调DKK3的表达。
3.3 miR-483-3p调控肿瘤发生相关基因
DKK3属于DKKs家族,一般认为它是信号分子Wnt的拮抗剂。DKK3可以通过与卷曲的受体结合来阻断Wnt信号通路,从而抑制Wnt/β-catenin信号通路的激活。我们的数据表明DKK3可能是miR483-3p的一个很有前途的靶基因,因此我们推测miR-483-3p可能影响结直肠癌Wnt/β-catenin信号通路,从而干扰结直肠癌的发展。沉默miR-483-3p可降低Wnt/β-catenin途径中的关键蛋白β-catenin的水平(图4A,B)。此外,转染miR-483-3p抑制剂的LoVo和SW480细胞c-Myc、cyclinD1、Bcl2水平表达降低,bax、E-cadherin水平表达增强,进一步提示miR-483-3p抑制剂可以抑制这两种细胞系Wnt/β-catenin蛋白通路的激活(图4A,B)。综上所述,DKK3-Wnt/β-catenin可能是miR-483-3p的直接靶点,提示miR-483-3p可能在体外调控肿瘤发生相关基因。
3.4 miR-483-3P对大肠癌细胞增殖和迁移的影响
Cyclin D1、CDK4、Bcl-2、Bax和E-cadherin是影响细胞迁移和增殖的基因。因此,为了进一步评价miR-483-3p的功能,将miR-483-3p抑制剂转染SW480和LoVo细胞,然后进行伤口愈合实验。转染miR-483-3p抑制剂后,LoVo和SW480细胞的迁移能力均下降(图4C,D)。此外,与未转染miR-483-3p抑制剂的细胞相比,这些细胞的活力明显降低(图5A,B)。在不同的实验中,用miR-483-3p抑制剂转染LoVo细胞和SW480细胞,进行集落形成实验。与未转染抑制剂的细胞相比,这些细胞的集落数量减少(图5C,D)。综上所述,结果提示miR-483-3p的下调可能会降低大肠癌细胞的增殖和迁移能力。
4 讨论
在本研究中发现,在AOM/DSS诱导的小鼠中,补充BRB花色苷膳食的小鼠miR-483-3p的表达显著降低。miR-483-3p可能通过调节Wnt/β-catenin信号通路,有效干扰癌细胞的迁移,从而发挥对大肠癌的治疗作用。
全世界都有证据表明,结直肠癌与高发病率和高死亡率有关。针对结直肠癌的治疗方法和结直肠癌患者的预后的进展仍然是不合需要的。因此,建立结直肠癌发病的潜在机制,寻找更有效的防治结直肠癌的方法具有重要意义。miRNAs是以相关mRNA的3‘-UTR为靶点、抑制其翻译的短链非编码RNA。miRNAs调控不同的生物学过程,包括细胞凋亡、细胞血管生成、细胞迁移和细胞增殖,通常通过靶向不同的mRNAs来表现为肿瘤抑制基因或癌基因。在本研究中,我们发现在CRC小鼠模型中给予BRB花色苷后,作为癌基因的miR-483-3p表达减少。肺癌、胰腺癌患者血浆中存在miR-483-3p的高表达;大肠癌中miR-483-3p也有高表达,但其具体作用机制尚未详细研究。大肠癌中检测到DKK3表达下调,生物信息学分析显示DKK3的表达可能影响患者的生存时间(图2C)。经BRB花色苷处理后,LoVo和SW480细胞的DKK3水平上调(图3g,H)。因此,BRB花色苷可以影响miR-483-3p和DKK3的表达。
DKK3已被证明可以抑制骨肉瘤、结肠癌、胃癌、胶质瘤、前列腺、宫颈癌、肝癌和肺癌中癌细胞的增殖。此外,DKK3也是Wnt信号通路的抑制剂,它参与了核β-catenin的降解,阻止了蛋白质向胞浆的转移。Wnt信号通路高度保守,在胚胎发育、动态平衡和肿瘤发生中发挥重要作用。Wnt/β-catenin信号通路参与细胞的侵袭、迁移、增殖和分化过程,在大肠癌的发生和发展中起重要作用。DKK3参与miR-483-3p介导的抑制CRC细胞增殖和迁移的作用,进一步证明了在用miR-483-3p特异性抑制剂处理时CRC细胞迁移和增殖能力降低(图4C、D和5A-D)。
以前,我们已经证明miR-24-1-5p的上调参与了BRB花色苷对CRC的化学预防,提示调节miRNA的表达可能是BRB花色苷对CRC发挥化学预防作用的一个可能机制。这也首次证明了miRNA可以参与BRB花色苷介导的抑制CRC的作用。本研究的发现进一步增强了miRNA在大肠癌发病机制中的重要性。它鉴定了另一个miRNA,miR-483-3p,证明了miR-483-3p在大肠癌发病机制中的潜在功能,并定位了其靶基因DKK3,作为揭示BRB花色苷抗癌活性介导的机制的一部分。因此,这项研究支持了我们的假设,即miRNAs参与了BRB花色苷介导的结直肠癌的化学预防。图6描绘了miR-483-3P可能通过BRB花色苷的化学预防作用来对抗结直肠癌的可能途径。我们通过miRNA芯片检测发现了受BRB花色苷调控的miRNAs。由于该方法的局限性以及小鼠组织样品与人组织样品的差异,我们认为miR-24-1-5p或miR-483-3p不是唯一受BRB花色苷调控的miRNA。然而,BRB花色苷对miRNA表达的调控仍不清楚。最近的报道描述了miRNA表达的调控是通过表观遗传机制进行的。在大肠癌和其他各种癌症中,miRNAs的表达受DNA甲基化的调节。例如,在HT-29 CRC细胞中miR-342的表达被高甲基化抑制。大肠癌中表观遗传沉默的miR-149与邻近CpG区域的高甲基化有关。许多参与DNA甲基化和组蛋白乙酰化调控的酶,如DNA甲基转移酶(DNMTs)、组蛋白脱乙酰酶或多梳抑制复合体基因等都参与了染色质结构的调控。有趣的是,许多研究表明,天然存在的生物活性化合物可以抑制这些酶的活性。例如,绿茶儿茶素中的活性多酚表没食子儿茶素-3-没食子酸酯可以抑制DNMTs的活性,从而抑制DNMT1、DNMT3a和DNMT3b的表达。此外,I期先导研究显示,黑树莓可以使人体内一些结直肠癌的抑癌基因去甲基化,而BRB花色苷也可以通过抑制结肠癌细胞中的DNMT1和DNMT3b使抑癌基因去甲基化。此外,在癌前病变组织中,黑树莓保护性地调节Wnt信号通路中基因的甲基化。因此,我们推测BRB花色苷对miRNA表达(以及可能的活性)的影响可能是通过BRB花色苷对表观遗传机制的影响来介导的。然而,BRB花色苷是否可以通过调节参与表观遗传过程的酶来影响miRNAs的表达,这还是一个有待进一步研究的课题。
综上所述,本研究结果表明,miR-483-3p可能通过其靶基因之一Dkk3调控Wnt/β-catenin途径在结直肠癌中发挥重要作用,并且miR-483-3p的表达可被BRB花色苷下调,进一步证明了BRB花色苷作为结直肠癌化学预防剂的潜力。此外,miR-483-3p的沉默可显著抑制大肠癌细胞的迁移和增殖,提示特异性靶向miR-483-3p的表达也可能成为未来治疗大肠癌的一种途径。
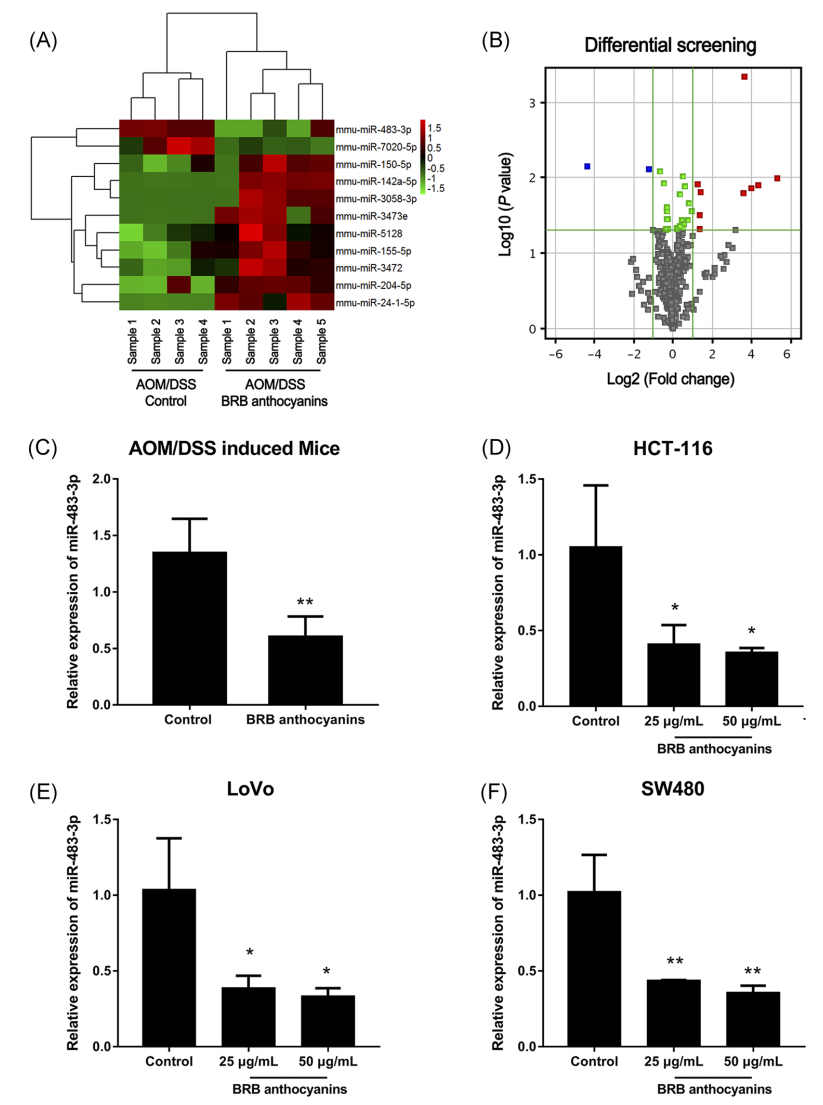
图1 BRB花色苷对AOM/DSS诱导小鼠及不同大肠癌细胞株miR-483-3p表达的影响。a、miRNAs在小鼠结肠中的差异表达。绿色-下调;红色-上调。B,下调的miRNAs用蓝色表示。定量RT-PCR检测AOM/DSS诱导的小鼠接受和不接受BRB花色苷后结肠组织miR-483-3p的水平。d、BRB花色苷处理前后HCT-116细胞miR-483-3p的相对表达。经BRB花色苷处理的LoVo细胞miR-483-3p的相对表达量。f,在加或不加BRB花色苷的SW480细胞中,miR-483-3p的相对表达。数据以均值±SEM表示。*和**分别在P<0.05和P<0.01水平上与对照组(不补充BRB)有显著差异。AOM,偶氮甲烷;BRB,黑莓;CRC,结直肠癌;DSS,葡聚糖硫酸钠;miRNAs,microRNAs;RT-PCR,实时聚合酶链反应[彩色图表可在wileyonlinelibrary.com查看]
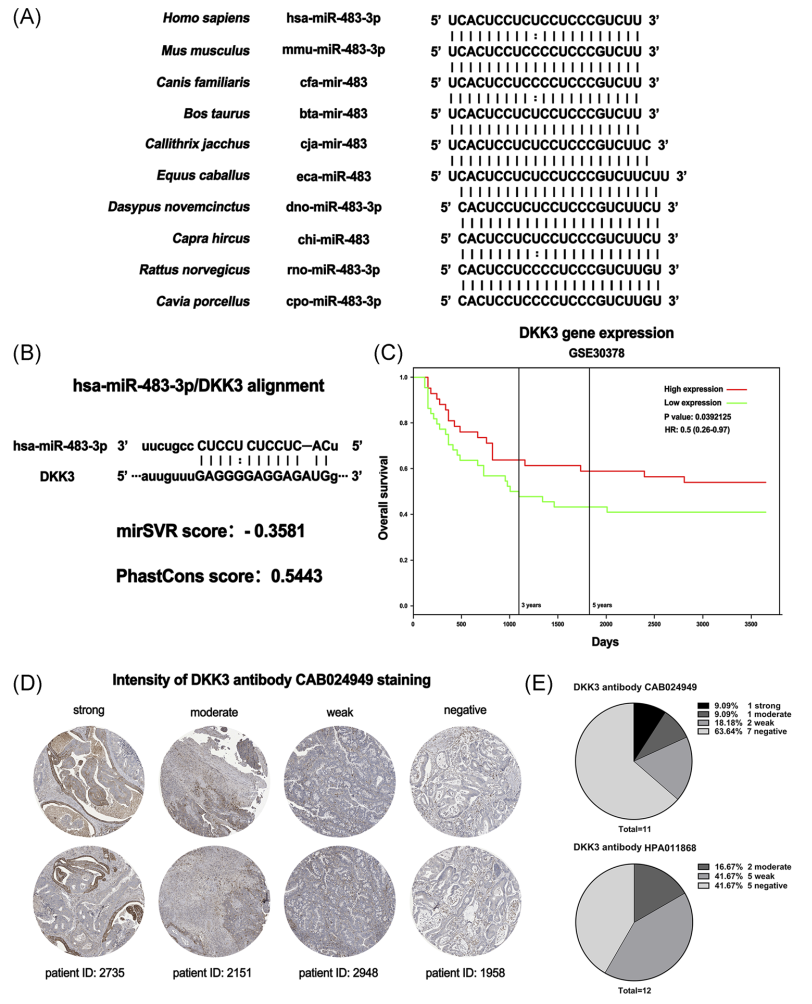
图2 利用生物信息学分析预测miR-483-3p靶基因。不同物种miR-483-3p序列的比对。B,靶向DKK3mRNA 3‘-UTR位点的miR-483-3p模型。根据在线数据库PROGeneV2的数据分析DKK3与大肠癌患者生存时间的关系。免疫组织化学染色显示大肠癌组织中D、DKK3水平。数据来源于人类蛋白质图谱数据库。图2D中DKK3蛋白表达的免疫组化染色数据统计分析。3‘-UTR,3’-非翻译区;CRC,结直肠癌;miR,microRNA[彩色图可在wileyonlinelibrary.com查看].
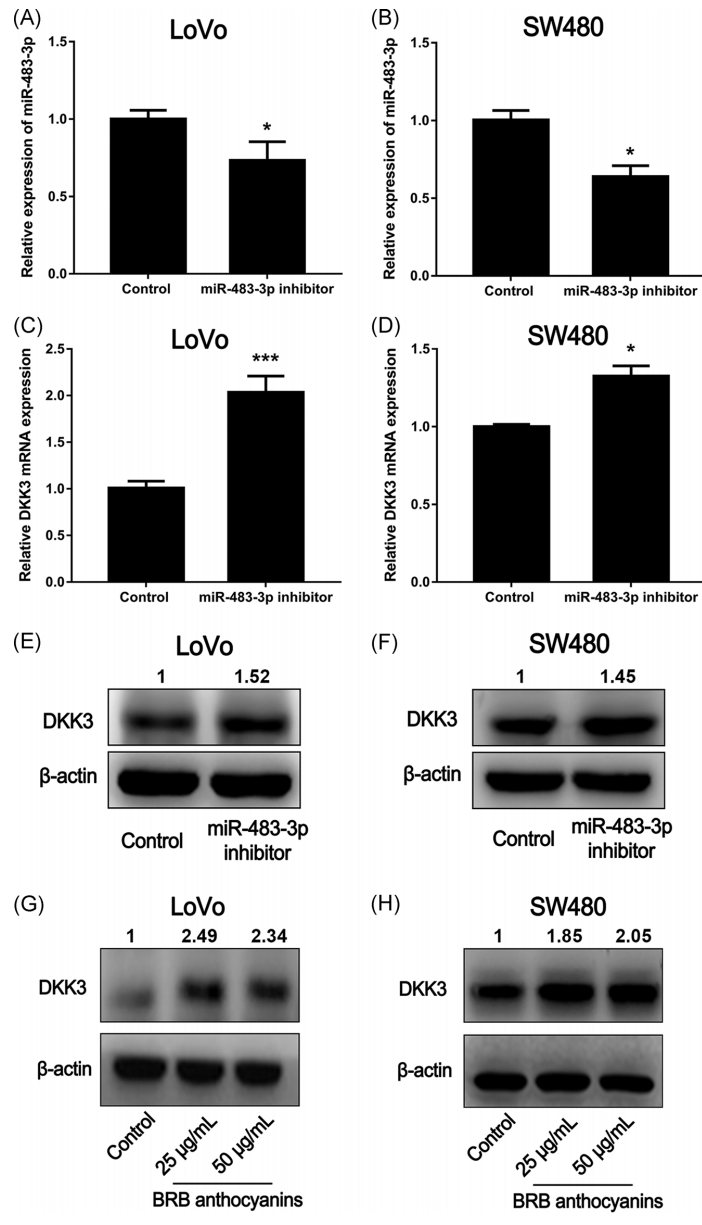
图3 miR-483-3p下调DKK3的表达。将miR-483-3p抑制剂转染LOVO和SW480细胞,qRT-PCR检测miR-483-3p(A和B)和DKK3(C和D)的表达水平,Western blot检测DKK3(E和F)的表达水平。Western blot检测BRB花色苷对LoVo(G)和SW480(H)细胞DKK3表达的影响。qRT-PCR数据为三次测定的平均值±SEMs。*和**分别与对照组(未加miR-483-3p抑制剂)在P<0.05和P<0.01水平上有显著性差异。对于Western blot分析,只显示有代表性的斑点。BRB,黑树莓;miR,microRNA;qRT-PCR,实时定量聚合酶链反应.
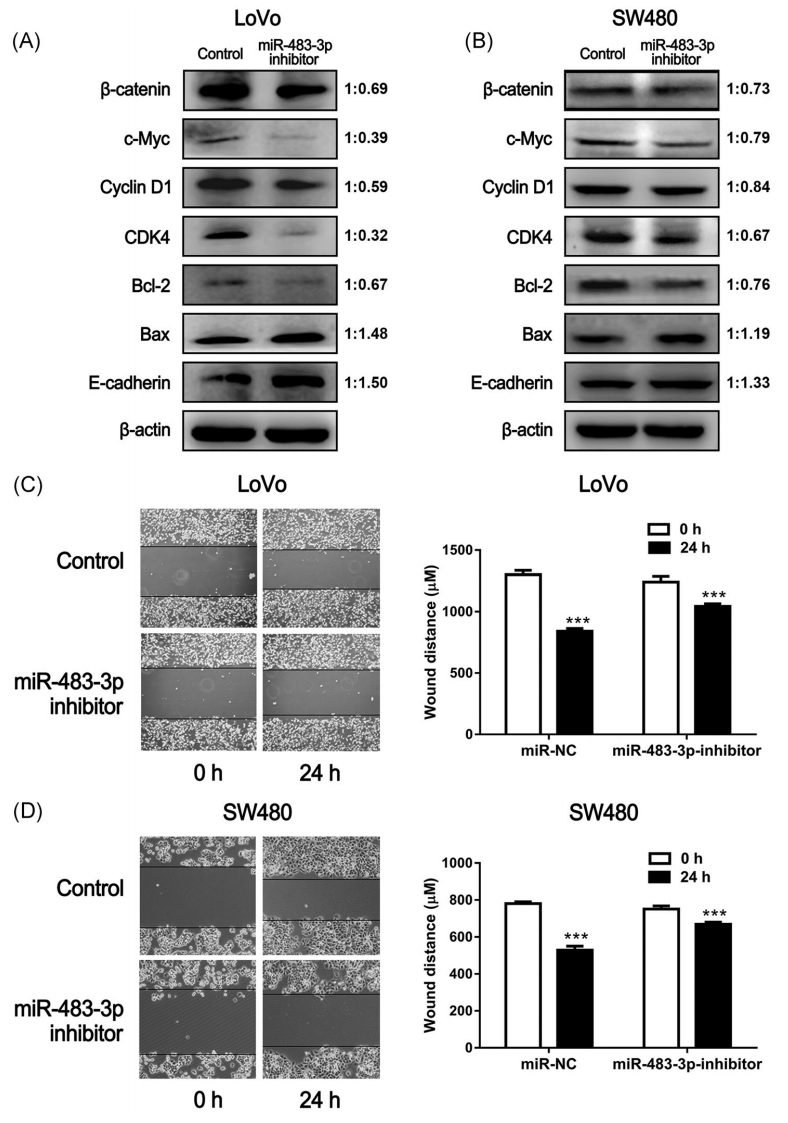
图4 miR-483-3p对肿瘤发生相关基因和细胞迁移的影响。Western blot分析转染β-483-3p抑制剂后LoVo(A)和SW480(B)细胞中c-myc、cyclinD1、CDK4、Bcl2、bax、E-cadherin和β-actin的表达水平。伤口愈合实验显示转染miR-483-3p抑制剂后的LoVo(C)和SW480(D)细胞迁移能力降低。图像旁边的图定量比较了在没有miR-483-p抑制剂和存在miR-483-p抑制剂的情况下相应细胞的迁移距离。数据以均值±SEM表示。*和**分别在P<0.05和P<0.01水平上与对照组(不补充BRB)有显著差异。BRB,黑色树莓;miR,microRNA。
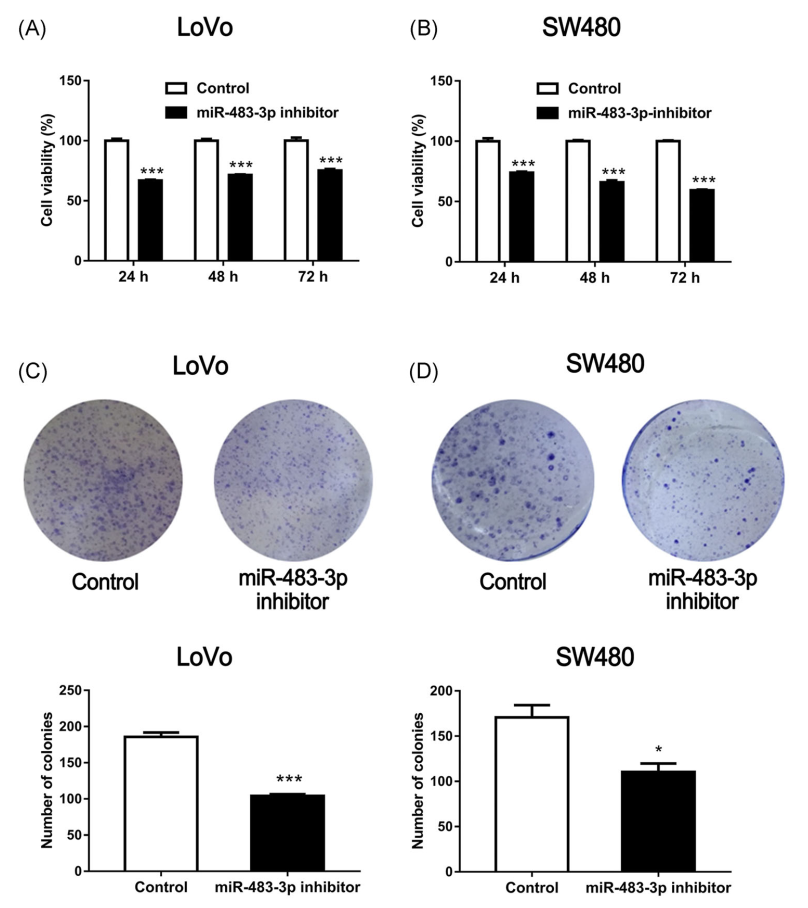
图5 miR-483-3p对大肠癌细胞增殖和集落形成的影响。将miR-483-3p抑制剂转染人大肠癌细胞、LoVo细胞和SW480细胞,A、B用MTT法检测细胞存活率,C、D用集落形成实验检测细胞增殖。这些曲线图显示了C和D中数据的定量比较。数据以均值±SEMS表示。*和**分别在P<0.05和P<0.01水平上与对照组(不补充BRB)有显著差异。BRB,黑色覆盆子;CRC,结直肠癌;MTT,3-(4,5-二甲基噻唑-2-基)-2,5-二苯基四唑溴[彩色图形可在wileyonlinelibrary.com查看]
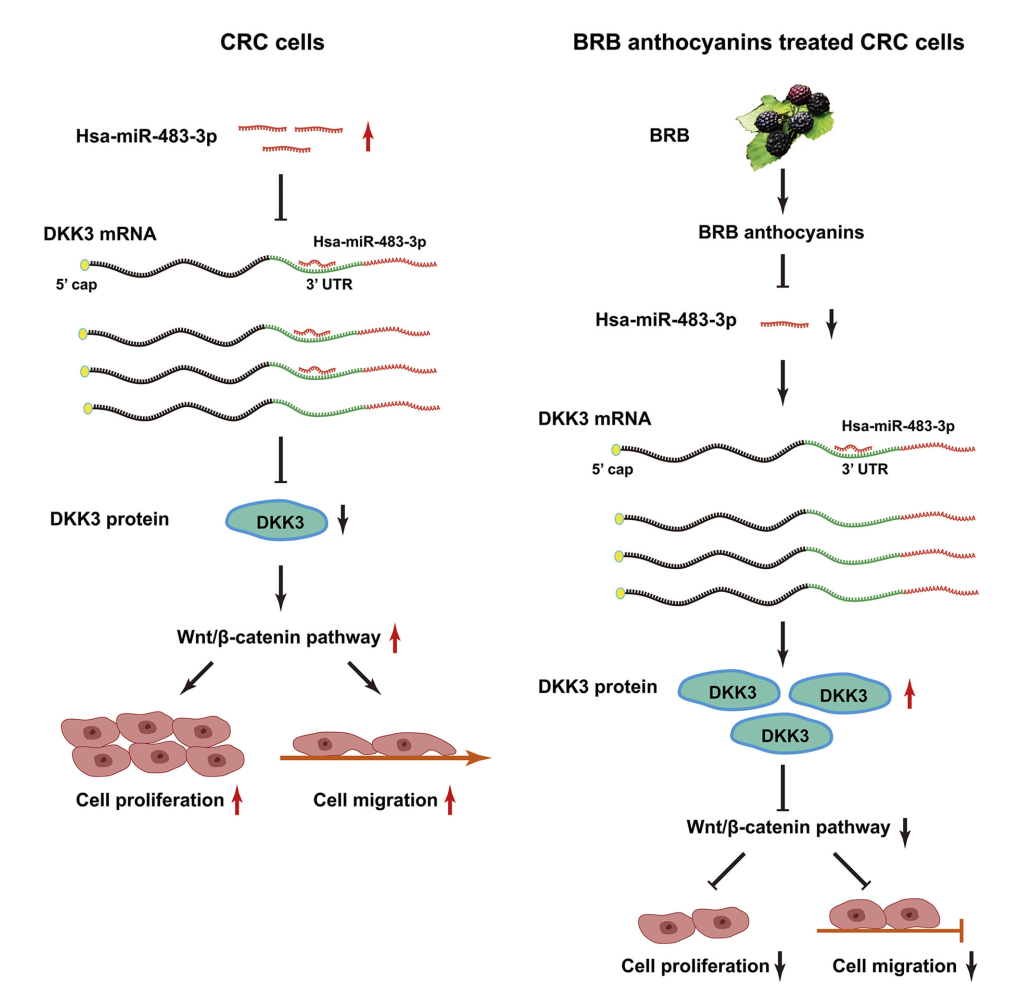
图6 这个模型描述了miR-483-3p如何通过BRB花青素的化学预防作用来对抗结直肠癌,在CRC细胞中,miR-483-3p可以通过下调DKK3的表达来促进细胞的增殖和迁移。BRB花色苷可抑制miR-483-3p,从而抑制大肠癌细胞的增殖和迁移。3‘-UTR,3’-非翻译区;BRB,黑莓;CRC,结直肠癌;miR,microRNA[彩色图形可在wileyonlinelibrary.com查看]
翻译:
miR-483-3P上调DKK3在黑树莓花色苷介导的结直肠癌化学预防中的重要作用
摘要:据报道,黑树莓(BRB)花色苷可作为一种潜在的结直肠癌(CRC)化学预防药物。然而,BRB花色苷抑制大肠癌细胞癌变的潜在机制尚未阐明。靶向重要抑癌基因的microRNAs(MiRNAs)的异常表达通常与大肠癌的发生有关。在本研究中,我们探讨了BRB花色苷在偶氮甲烷(AOM)/葡聚糖硫酸钠(DSS)诱导的小鼠大肠癌模型和人大肠癌细胞系中是否影响某些miRNAs的表达。采用miRNA微阵列分析方法,研究饲喂不含或不含BRB花青素的饲料诱导的AOM/DSS小鼠miRNA表达的差异。一种特殊的miRNA,miR-483-3p的表达在AOM/DSS诱导的小鼠中被发现在饲料中添加BRB花青素后显著降低。随后的实时定量聚合酶链反应和Western blot分析表明,miR-483-3p表达降低的同时,生物信息学分析预测的miR-483-3p的潜在靶点Dickkopf3(DKK3)的表达增加。当miR-483-3p特异性抑制剂下调miR-483-3p的表达水平时,DKK3的蛋白和信使RNA水平明显上调,提示DKK3可能是miR-483-3p的靶基因。此外,包括Wnt/β-catenin在内的DKK3信号通路下游因子也在MIR483-3p介导的BRB花色苷抗癌作用中发挥作用。因此,miR-483-3p可能是BRB花色苷介导的预防大肠癌的潜在靶点。
关键词:
BRB花色苷、结直肠癌、DKK3、miR-483-3p、Wnt/β-catenin通路
1、简介
MicroRNAs(MiRNAs)是一种小分子单链RNA,普遍存在于动植物中。它通过与靶基因mRNA的3’-非翻译区(3’-UTR)结合,抑制翻译或直接降解信使RNA(MRNA),直接影响细胞生长、器官形成、能量代谢、细胞增殖、凋亡和防御系统。miRNAs还参与癌症的发展,既可以是癌基因,也可以是抑癌基因。例如,miR-21-5p和miR-142-5p既可以作为致癌基因也可以作为抑癌基因,而miR-22、miR-490-3p和miR-93都可以作为肿瘤抑制基因。此外,miRNAs还可以作为检测癌症的有用标记物。例如,血液中的miR-193a-3p、miR-23a和miR-338-5p可用于大肠癌(CRC)的早期检测。miR-483-3p通过抑制DPC4/Smad4的表达发挥促癌作用,从而促进胰腺癌的细胞增殖和克隆性。随着越来越多的miRNAs被发现在结直肠癌中异常表达,miRNA的应用将成为结直肠癌诊断、靶向治疗和预后研究的热点。大肠癌是一种常见的癌症,是世界上第四大常见死亡原因。大肠癌的发生发展涉及多个基因和信号通路的改变。大肠癌的发生发展伴随着Wnt/β-catenin信号通路的异常激活。Dickkopf蛋白家族由DKK1、DKK2、DKK3和DKK4组成,是Wnt/β-catenin信号通路的细胞外抑制因子,它通过与LRP5/LRP5受体相互作用来影响Wnt信号通路的非经典激活。
长期食用黑树莓(BRB)可以降低患癌症、心脏病、关节炎和呼吸系统疾病的风险。食用BRB会产生更多的次生代谢物,从而改善健康。BRB花色苷是BRB在体内产生的主要代谢物之一,也是膳食中花色苷的主要来源。BRB最显著的有益作用是其潜在的癌症化学预防作用,已有多项研究对此进行了研究。我们先前的研究发现,BRB花色苷可以调节肠道微生物区系,进一步使分泌的卷曲相关蛋白5的启动子去甲基化,最终延缓偶氮甲烷/葡聚糖硫酸钠(AOM/DSS)处理小鼠的结直肠癌的发展。花色苷似乎是临床试验中使用的冷冻干燥BRB粉末中的关键抗癌成分。
我们最近的研究发现miR-24-1-5p可以下调Wnt/β-catenin信号通路,有助于其在预防大肠癌的发生发展中发挥作用,从而参与BRB花色苷介导的结直肠癌的化学预防过程中的miRNAs的作用。在本研究中,miR-24-1-5p可下调Wnt/miRNAs-catenin信号通路的表达,从而起到预防大肠癌发生的作用。在这项研究中,我们研究了另一种miRNA miR-483-3p的作用,以确定它可能通过口服BRB花色苷抑制AOM/DSS诱导的CRC小鼠模型中CRC的发展的机制。在用BRB花色苷处理人大肠癌细胞后,还检测了miR-483-3p的作用。
2、材料和方法
2.1收购BRB花青素
BRBs含有各种化学物质,如酚酸和花色苷。BRB花色苷的主要成分为花青素-3-O-葡萄糖苷、花青素-3-O-芦丁苷、花青素-3-O-木糖基-芦丁苷等。本研究采用JF Natural公司(天津建峰天然产品研发有限公司)提取BRB花色苷。简单地说,冷冻干燥的BRB粉末用乙醇-水(75:25,体积/体积)在35°C下搅拌3h,直至粉末变白,得到粗提物。萃取物经滤纸过滤后装入聚酰胺色谱柱分离成不同组分。进一步用制备高效液相色谱分离纯化得到纯化的BRB花色苷。BRB花色苷保存在−20°C的环境中。
2.2 CRC小鼠模型的建立
将20只C57雄性小鼠(体重18~20g,辽宁长生实验动物生物技术有限公司)随机分为2组,每组10只。其中一组被指定为模型组,按正常饮食喂养。另一组为试验组,饲喂含7.0molmol/g BRB花色苷的普通饲料。饲料中BRB花色苷的浓度是在我们前期研究的基础上选择的。两组小鼠在实验的第一天注射AOM(10 mg/kg),然后在实验的第8天再注射。同时,第1周每天用2%DSS水治疗,随后2周每天用正常水治疗,整个过程重复3次。因此,整个实验持续了9周,在治疗期结束时,处死并取出它们的结肠,并将其冷冻在液氮中。部分组织用于miRNA阵列分析,其余组织保存在−80°C进行进一步分析。
在整个实验过程中,小鼠被关在笼子里(每笼3~5只),保持在22°C±1°C和50%±5%的湿度下,光照-黑暗周期为12小时(上午7:00-晚上7:00)。所有的程序都是根据国家卫生研究院关于动物护理和使用的规定进行的,并得到了辽宁中医药大学伦理委员会的批准。
2.3 miRNA微阵列
提取的结肠组织进行微阵列分析。使用安捷伦(安捷伦,Palo Alto,CA)对原始数据中不同基因的表达进行微阵列(8*60K,设计ID:070155)和基本分析。
2.4 细胞培养
人HCT116和HT29大肠癌细胞系购自中国科学院(上海)细胞库。LOVO细胞和SW480细胞购自上海明景生物研究所。细胞培养在37°C的5%CO2加湿环境中。
2.5 miRNAs瞬时转染
miR-483-3P抑制剂和miR-NC购自GenePharma(中国上海)。LoVo和SW480细胞的瞬时转染按照制造商方案进行。
2.6 RNA提取及qRT-PCR
用TRIzol试剂(上海桑贡生物科技有限公司)从细胞和组织中提取总RNA。编码miRNA的第一链互补DNA(cDNA)用miRNA cDNA合成试剂盒(ComWin Biotech Co,Ltd,China,China)反转录获得。用ComWin Biotech Co,Ltd的miRNA实时检测试剂盒进行miR-483-3p的实时定量聚合酶链反应(qRT-PCR)。用PrimeScript RT试剂盒检测DKK3的mRNA水平,试剂盒采用gDNA擦除器和TB Green PreMix Ex Taq II(北京塔卡拉生物医学技术有限公司)。用特异引物(表S1)进行qRT-PCR,并使用应用生物系统7500实时PCR系统进行扩增。
2.7 蛋白质提取及Western blot分析
冰冻条件下先用放射免疫沉淀分析缓冲液(北京鼎国长生生物科技有限公司)裂解细胞,随后细胞裂解液离心。然后,使用针对DKK3、β-肌动蛋白、Bcl-2、BAX(Sangon Biotech Shanghai Co,Ltd)、Cyclin D1、c-Myc、CDK4或E-cadherin(细胞信号技术,MA)的抗体,对所获得的含有可溶性蛋白的上清液进行Western blot分析。使用Image J软件对条带强度进行量化。
2.8 伤口愈合试验
这些细胞在一个六孔培养板中培养,直到它们达到60%的融合。然后用无菌微吸管刮除细胞层形成伤口,并用新鲜培养基更换培养液。然后,将miRNAs转染细胞24小时,拍摄细胞层的显微照片。光镜下计算细胞迁移程度。
2.9 3-(4,5-二甲基噻唑-2-基)-2,5-二苯基四唑溴化法
BRB花色苷的原液用培养基稀释到合适的浓度,然后加入96孔板,其中包含生长到60%融合的细胞。在细胞中分别加入BRB花色苷到最终浓度为25和50μg/mL。对照组仅向细胞中加入等量培养基,而不加入BRB花色苷。用3-(4,5-二甲基噻唑-2-基)-2,5-二苯基四唑溴化物(Sangon Biotech Shanghai Co,Ltd)试剂(Sangon Biotech Shanghai Co,Ltd)测定细胞增殖。
2.10 miRNA靶基因的鉴定及其与大肠癌患者生存率的关系
利用在线数据库microRNA.org预测miR-483-3p的靶基因(http://www.microrna.org/microrna/home.do)。从人类蛋白质图谱网站(www.proteinatlas.org)获得结直肠癌患者结肠组织中DKK3的表达谱。并用miRbase(http://www.mirbase.org/)对不同物种的miR-483-3p序列进行比对。使用在线数据库PROGeneV2(http://genomics.jefferson.edu/proggene/)验证DKK3的表达与大肠癌患者生存率的关系。
2.11 统计方式
采用SPSS 19.0统计软件进行数据分析。不同组间比较采用t检验和方差分析。在P<0.05和P<0.01水平上有统计学意义。
3 结果
3.1 miR-483-3p可能在BRB花色苷介导的大肠癌预防中发挥作用
应用miRNA微阵列分析方法获得了AOM/DSS诱导的小鼠结肠中miRNA的表达特征,这些小鼠分别饲喂了不添加(对照)或添加BRB花色苷的饲料。在接受BRB花色苷组中,几个miRNAs的表达模式与对照组不同。接受BRB花色苷的小鼠显示其结肠中miR-483-3p的表达减少(图1A,B)。此外,qRT-PCR显示,经BRB花色苷处理的CRC细胞与未处理的CRC细胞相比,miR-483-3p的表达也降低(图1C和1F)。提示miR-483-3p可能在BRB花色苷介导的化学预防大肠癌中起作用。
3.2 DKK3是miR-483-3p的预测靶基因,DKK3的表达水平与大肠癌患者有关
为了更好地了解miR-483-3p在大肠癌中的意义,我们对不同物种的miR-483序列进行了比对。结果显示,高等哺乳动物的miR-483序列具有高度的保守性(图2A)。此外,生物信息学分析预测DKK3可能是miR-483-3p的潜在靶基因(图2B)。然后利用在线数据库PROGeneV2分析DKK3与大肠癌患者生存时间的关系。DKK3 mRNA水平与患者总生存率呈正相关(图2C)。此外,从人类蛋白图谱数据库网站获得的DKK3蛋白免疫组织化学染色数据显示,这些大肠癌患者中DKK3的表达下调(图2D,E)。综上所述,DKK3可能是miR-483-3p在大肠癌患者中低表达的靶基因。BRB花色苷可能通过下调miR-483-3p的表达来调节DKK3的表达。
为了进一步检测miR-483-3p对DKK3的作用,用miR-483-3p抑制剂转染LoVo细胞和SW480细胞,该抑制剂由与miR-483-3p序列互补的寡核苷酸组成。转染miR-483-3p抑制剂后,细胞中DKK3的蛋白和mRNA水平均显著升高(图3A-C和3F)。此外,经BRB花色苷处理的LoVo和SW480细胞显示DKK3蛋白水平升高(图3g,H),进一步支持DKK3可能是miR-483-3p的靶基因的推测,BRB花色苷可能通过下调miR-483-3p表达来下调DKK3的表达。
3.3 miR-483-3p调控肿瘤发生相关基因
DKK3属于DKKs家族,一般认为它是信号分子Wnt的拮抗剂。DKK3可以通过与卷曲的受体结合来阻断Wnt信号通路,从而抑制Wnt/β-catenin信号通路的激活。我们的数据表明DKK3可能是miR483-3p的一个很有前途的靶基因,因此我们推测miR-483-3p可能影响结直肠癌Wnt/β-catenin信号通路,从而干扰结直肠癌的发展。沉默miR-483-3p可降低Wnt/β-catenin途径中的关键蛋白β-catenin的水平(图4A,B)。此外,转染miR-483-3p抑制剂的LoVo和SW480细胞c-Myc、cyclinD1、Bcl2水平表达降低,bax、E-cadherin水平表达增强,进一步提示miR-483-3p抑制剂可以抑制这两种细胞系Wnt/β-catenin蛋白通路的激活(图4A,B)。综上所述,DKK3-Wnt/β-catenin可能是miR-483-3p的直接靶点,提示miR-483-3p可能在体外调控肿瘤发生相关基因。
3.4 miR-483-3P对大肠癌细胞增殖和迁移的影响
Cyclin D1、CDK4、Bcl-2、Bax和E-cadherin是影响细胞迁移和增殖的基因。因此,为了进一步评价miR-483-3p的功能,将miR-483-3p抑制剂转染SW480和LoVo细胞,然后进行伤口愈合实验。转染miR-483-3p抑制剂后,LoVo和SW480细胞的迁移能力均下降(图4C,D)。此外,与未转染miR-483-3p抑制剂的细胞相比,这些细胞的活力明显降低(图5A,B)。在不同的实验中,用miR-483-3p抑制剂转染LoVo细胞和SW480细胞,进行集落形成实验。与未转染抑制剂的细胞相比,这些细胞的集落数量减少(图5C,D)。综上所述,结果提示miR-483-3p的下调可能会降低大肠癌细胞的增殖和迁移能力。
4 讨论
在本研究中发现,在AOM/DSS诱导的小鼠中,补充BRB花色苷膳食的小鼠miR-483-3p的表达显著降低。miR-483-3p可能通过调节Wnt/β-catenin信号通路,有效干扰癌细胞的迁移,从而发挥对大肠癌的治疗作用。
全世界都有证据表明,结直肠癌与高发病率和高死亡率有关。针对结直肠癌的治疗方法和结直肠癌患者的预后的进展仍然是不合需要的。因此,建立结直肠癌发病的潜在机制,寻找更有效的防治结直肠癌的方法具有重要意义。miRNAs是以相关mRNA的3‘-UTR为靶点、抑制其翻译的短链非编码RNA。miRNAs调控不同的生物学过程,包括细胞凋亡、细胞血管生成、细胞迁移和细胞增殖,通常通过靶向不同的mRNAs来表现为肿瘤抑制基因或癌基因。在本研究中,我们发现在CRC小鼠模型中给予BRB花色苷后,作为癌基因的miR-483-3p表达减少。肺癌、胰腺癌患者血浆中存在miR-483-3p的高表达;大肠癌中miR-483-3p也有高表达,但其具体作用机制尚未详细研究。大肠癌中检测到DKK3表达下调,生物信息学分析显示DKK3的表达可能影响患者的生存时间(图2C)。经BRB花色苷处理后,LoVo和SW480细胞的DKK3水平上调(图3g,H)。因此,BRB花色苷可以影响miR-483-3p和DKK3的表达。
DKK3已被证明可以抑制骨肉瘤、结肠癌、胃癌、胶质瘤、前列腺、宫颈癌、肝癌和肺癌中癌细胞的增殖。此外,DKK3也是Wnt信号通路的抑制剂,它参与了核β-catenin的降解,阻止了蛋白质向胞浆的转移。Wnt信号通路高度保守,在胚胎发育、动态平衡和肿瘤发生中发挥重要作用。Wnt/β-catenin信号通路参与细胞的侵袭、迁移、增殖和分化过程,在大肠癌的发生和发展中起重要作用。DKK3参与miR-483-3p介导的抑制CRC细胞增殖和迁移的作用,进一步证明了在用miR-483-3p特异性抑制剂处理时CRC细胞迁移和增殖能力降低(图4C、D和5A-D)。
以前,我们已经证明miR-24-1-5p的上调参与了BRB花色苷对CRC的化学预防,提示调节miRNA的表达可能是BRB花色苷对CRC发挥化学预防作用的一个可能机制。这也首次证明了miRNA可以参与BRB花色苷介导的抑制CRC的作用。本研究的发现进一步增强了miRNA在大肠癌发病机制中的重要性。它鉴定了另一个miRNA,miR-483-3p,证明了miR-483-3p在大肠癌发病机制中的潜在功能,并定位了其靶基因DKK3,作为揭示BRB花色苷抗癌活性介导的机制的一部分。因此,这项研究支持了我们的假设,即miRNAs参与了BRB花色苷介导的结直肠癌的化学预防。图6描绘了miR-483-3P可能通过BRB花色苷的化学预防作用来对抗结直肠癌的可能途径。我们通过miRNA芯片检测发现了受BRB花色苷调控的miRNAs。由于该方法的局限性以及小鼠组织样品与人组织样品的差异,我们认为miR-24-1-5p或miR-483-3p不是唯一受BRB花色苷调控的miRNA。然而,BRB花色苷对miRNA表达的调控仍不清楚。最近的报道描述了miRNA表达的调控是通过表观遗传机制进行的。在大肠癌和其他各种癌症中,miRNAs的表达受DNA甲基化的调节。例如,在HT-29 CRC细胞中miR-342的表达被高甲基化抑制。大肠癌中表观遗传沉默的miR-149与邻近CpG区域的高甲基化有关。许多参与DNA甲基化和组蛋白乙酰化调控的酶,如DNA甲基转移酶(DNMTs)、组蛋白脱乙酰酶或多梳抑制复合体基因等都参与了染色质结构的调控。有趣的是,许多研究表明,天然存在的生物活性化合物可以抑制这些酶的活性。例如,绿茶儿茶素中的活性多酚表没食子儿茶素-3-没食子酸酯可以抑制DNMTs的活性,从而抑制DNMT1、DNMT3a和DNMT3b的表达。此外,I期先导研究显示,黑树莓可以使人体内一些结直肠癌的抑癌基因去甲基化,而BRB花色苷也可以通过抑制结肠癌细胞中的DNMT1和DNMT3b使抑癌基因去甲基化。此外,在癌前病变组织中,黑树莓保护性地调节Wnt信号通路中基因的甲基化。因此,我们推测BRB花色苷对miRNA表达(以及可能的活性)的影响可能是通过BRB花色苷对表观遗传机制的影响来介导的。然而,BRB花色苷是否可以通过调节参与表观遗传过程的酶来影响miRNAs的表达,这还是一个有待进一步研究的课题。
综上所述,本研究结果表明,miR-483-3p可能通过其靶基因之一Dkk3调控Wnt/β-catenin途径在结直肠癌中发挥重要作用,并且miR-483-3p的表达可被BRB花色苷下调,进一步证明了BRB花色苷作为结直肠癌化学预防剂的潜力。此外,miR-483-3p的沉默可显著抑制大肠癌细胞的迁移和增殖,提示特异性靶向miR-483-3p的表达也可能成为未来治疗大肠癌的一种途径。
图1 BRB花色苷对AOM/DSS诱导小鼠及不同大肠癌细胞株miR-483-3p表达的影响。a、miRNAs在小鼠结肠中的差异表达。绿色-下调;红色-上调。B,下调的miRNAs用蓝色表示。定量RT-PCR检测AOM/DSS诱导的小鼠接受和不接受BRB花色苷后结肠组织miR-483-3p的水平。d、BRB花色苷处理前后HCT-116细胞miR-483-3p的相对表达。经BRB花色苷处理的LoVo细胞miR-483-3p的相对表达量。f,在加或不加BRB花色苷的SW480细胞中,miR-483-3p的相对表达。数据以均值±SEM表示。*和**分别在P<0.05和P<0.01水平上与对照组(不补充BRB)有显著差异。AOM,偶氮甲烷;BRB,黑莓;CRC,结直肠癌;DSS,葡聚糖硫酸钠;miRNAs,microRNAs;RT-PCR,实时聚合酶链反应[彩色图表可在wileyonlinelibrary.com查看]
图2 利用生物信息学分析预测miR-483-3p靶基因。不同物种miR-483-3p序列的比对。B,靶向DKK3mRNA 3‘-UTR位点的miR-483-3p模型。根据在线数据库PROGeneV2的数据分析DKK3与大肠癌患者生存时间的关系。免疫组织化学染色显示大肠癌组织中D、DKK3水平。数据来源于人类蛋白质图谱数据库。图2D中DKK3蛋白表达的免疫组化染色数据统计分析。3‘-UTR,3’-非翻译区;CRC,结直肠癌;miR,microRNA[彩色图可在wileyonlinelibrary.com查看].
图3 miR-483-3p下调DKK3的表达。将miR-483-3p抑制剂转染LOVO和SW480细胞,qRT-PCR检测miR-483-3p(A和B)和DKK3(C和D)的表达水平,Western blot检测DKK3(E和F)的表达水平。Western blot检测BRB花色苷对LoVo(G)和SW480(H)细胞DKK3表达的影响。qRT-PCR数据为三次测定的平均值±SEMs。*和**分别与对照组(未加miR-483-3p抑制剂)在P<0.05和P<0.01水平上有显著性差异。对于Western blot分析,只显示有代表性的斑点。BRB,黑树莓;miR,microRNA;qRT-PCR,实时定量聚合酶链反应.
图4 miR-483-3p对肿瘤发生相关基因和细胞迁移的影响。Western blot分析转染β-483-3p抑制剂后LoVo(A)和SW480(B)细胞中c-myc、cyclinD1、CDK4、Bcl2、bax、E-cadherin和β-actin的表达水平。伤口愈合实验显示转染miR-483-3p抑制剂后的LoVo(C)和SW480(D)细胞迁移能力降低。图像旁边的图定量比较了在没有miR-483-p抑制剂和存在miR-483-p抑制剂的情况下相应细胞的迁移距离。数据以均值±SEM表示。*和**分别在P<0.05和P<0.01水平上与对照组(不补充BRB)有显著差异。BRB,黑色树莓;miR,microRNA。
图5 miR-483-3p对大肠癌细胞增殖和集落形成的影响。将miR-483-3p抑制剂转染人大肠癌细胞、LoVo细胞和SW480细胞,A、B用MTT法检测细胞存活率,C、D用集落形成实验检测细胞增殖。这些曲线图显示了C和D中数据的定量比较。数据以均值±SEMS表示。*和**分别在P<0.05和P<0.01水平上与对照组(不补充BRB)有显著差异。BRB,黑色覆盆子;CRC,结直肠癌;MTT,3-(4,5-二甲基噻唑-2-基)-2,5-二苯基四唑溴[彩色图形可在wileyonlinelibrary.com查看]
图6 这个模型描述了miR-483-3p如何通过BRB花青素的化学预防作用来对抗结直肠癌,在CRC细胞中,miR-483-3p可以通过下调DKK3的表达来促进细胞的增殖和迁移。BRB花色苷可抑制miR-483-3p,从而抑制大肠癌细胞的增殖和迁移。3‘-UTR,3’-非翻译区;BRB,黑莓;CRC,结直肠癌;miR,microRNA[彩色图形可在wileyonlinelibrary.com查看]
本文由福山生物整理翻译,转载请注明出处。