






最新资讯



-
电话: 400-113-6988 邮箱: dongfangxicao@163.com 地址: 深圳市南山区粤海街道高新中三道9号环球数码大厦19楼
↓ 微信扫码识别关注我们 ↓ 
SFN通过降低AGEs受体表达,抑制AGEs诱导的细胞损伤
发表于:2020-04-28 作者:admin 来源:本站 点击量:10665
Sulforaphane inhibits advanced glycation end product–induced pericyte damage by reducing expression of receptor for advanced glycation end products
SFN通过降低晚期糖基化终产物受体的表达,抑制晚期糖基化终产物诱导的周细胞损伤
晚期糖基化终产物(AGEs)不仅抑制DNA合成,而且还通过与AGE受体(RAGE)相互作用引起视网膜周细胞中的细胞凋亡和炎症,从而在糖尿病性视网膜病变中发挥作用。类似地,萝卜硫素是在广泛食用的十字花科蔬菜中发现的天然存在的异硫氰酸酯,其保护免受氧化应激诱导的组织损伤。因此,我们假设萝卜硫素可通过其抗氧化特性抑制AGE诱导的周细胞损伤。晚期糖基化终产物刺激了牛培养的视网膜周细胞中超氧化物的产生以及RAGE基因和蛋白质的表达,并且这些效果通过用萝卜硫素处理来防止。针对RAGE的萝卜硫素和抗体显著抑制了AGE诱导的DNA合成下降、凋亡细胞死亡以及周细胞单核细胞趋化蛋白1信使RNA水平的上调。本研究首次表明,萝卜硫素可抑制AGE暴露周细胞的DNA合成,凋亡细胞死亡和炎症反应,部分原因是通过其抗氧化特性抑制RAGE表达。通过萝卜硫素阻断周细胞中的AGE-RAGE轴可能是治疗糖尿病性视网膜病变的新型治疗靶点。
1. 引言
糖尿病视网膜病变是糖尿病中最具破坏性的微血管并发症之一,也是职业年龄人群获得性失明的主要原因[1,2]。糖尿病视网膜病变最早的组织病理学标志是周细胞丢失[3]。与周细胞损失同时,观察到几种特征性变化,包括视网膜基底膜增厚,视网膜血管通透性过高和微动脉瘤形成[3]。由于周细胞已被证明在维持微血管稳态中起着重要作用,因此推测糖尿病视网膜病变的恶化和与视网膜新生血管形成相关的增殖性变化是周细胞丢失的结果[4-6]。
晚期糖基化终产物(AGEs)由美拉德过程形成,这是一种还原糖和蛋白质氨基之间的非酶促反应,有助于蛋白质的老化[6-10]。在糖尿病高血糖条件下,这一过程开始于可逆的希夫碱加合物转化为更稳定的阿马多里(Amadori)产物。在数天到数周的过程中,Amadori产物经历进一步的重排反应,例如脱水和缩合,以形成称为AGE的不可逆交联的大蛋白衍生物。有越来越多的证据表明,AGE及其细胞表面受体,AGE相互作用受体(RAGE)与许多糖尿病和衰老相关疾病的病理后遗症的发展和进展有关[6-10]。事实上,我们以前发现AGEs不仅抑制DNA合成,而且还通过与RAGE的相互作用,通过氧化应激的产生诱导培养的视网膜周细胞中的凋亡细胞死亡[6,11,12]。AGE相互作用的晚期糖基化终产物受体也刺激各种细胞类型中的单核细胞趋化蛋白1(MCP-1)表达,这可能导致血管通透性过高和早期糖尿病视网膜病变血液 - 视网膜屏障功能受损[13-17]。这些观察结果表明,阻断AGE诱导的周细胞损伤可能是糖尿病视网膜病变的新治疗靶点。
萝卜硫素是十字花科蔬菜中天然存在的异硫氰酸酯,如西兰花、卷心菜和球芽甘蓝[18,19]。萝卜硫素是具有抗癌特性的II相抗氧化解毒酶的诱导剂[18,19]。此外,萝卜硫素对氧化应激相关组织损伤具有保护作用,包括实验性糖尿病肾病和神经病变[20,21]。萝卜硫素还被证明可以改善1型糖尿病大鼠的代谢紊乱,并通过Nrf2[20]的激活抑制氧化应激的产生,从而抑制蛋白尿和肾小球硬化。尽管已知萝卜硫素的许多有益属性,但据我们所知,尚无研究检测萝卜硫素对AGE诱发的周细胞损伤的影响。因此,我们假设萝卜硫素通过其抗氧化特性可以预防AGE-RAGE诱导的周细胞损伤。为了验证这一假设,我们研究了萝卜硫素是否能抑制AGE暴露的周细胞中超氧化物的产生,RAGE表达,DNA合成,细胞凋亡和炎症反应。
2. 方法和材料
2.1. 材料
D-甘油醛和1%蛋白酶抑制剂混合物购自Nakalai Tesque(Kyoto,Japan),[3H]胸苷购自GE Healthcare(Buckinghamshire,UK)。4-(2-羟乙基)-1-哌嗪乙磺酸和氯化钠来自日本大阪的Wako Pure Chemical Industries,Ltd。其他化学品购自Sigma(St Louis,MO,USA)。
2.2. AGE-牛血清白蛋白的制备
如前所述制备晚期糖基化终产物 - 牛血清白蛋白(BSA)[12]。简言之,将BSA(25mg / mL)在无菌条件下与0.1mol / L甘油醛在0.2mol / L磷酸钠缓冲液(pH7.4)中培育7天。然后,通过PD-10柱层析除去未掺入的糖并用磷酸盐缓冲盐水透析。除不含还原糖外,对照组未糖化BSA在相同条件下培育。使用Endospecy ES-20S系统(Seikagaku Co,Tokyo,Japan)测试制剂的内毒素; 没有检测到内毒素。
2.3. 制备针对RAGE的抗体
如前所述,制备了针对RAGE的抗体(RAGE-Ab),用于中和测定,其识别人RAGE蛋白的氨基酸残基167-180 [22]。
2.4. 细胞
分离牛视网膜周细胞并将其保持在Dulbecco 改良的Eagle培养基(Gibco BRL,Rockville,MD,USA)中,其如前所述[21,23]补充有20%胎牛血清(ICN Biomedicals Inc,Aurora,OH,USA)。将5至10次传代的细胞用于本实验。晚期糖基化终产物治疗在含有2%胎牛血清的培养基中进行。
2.5. 活性氧生成
在存在或不存在0.1或0.4μmol/ L萝卜硫素或5μg/ mL RAGE-Ab的情况下,用100μg/ mL AGE-BSA或非糖基化的BSA处理周细胞2小时。接下来,如前所述测量细胞内活性氧(ROS)的产生[24]。简而言之,在用磷酸盐缓冲盐水洗涤除去死细胞后,将周细胞在37℃下与不含酚红的Dulbecco改良的Eagle培养基一起培养15分钟,其中含有10μmol/ L 5-(和-6) - 氯甲基 -2',7'-二氯二氢荧光素二乙酸酯(Molecular Probes Inc,Eugene,OR,USA)。根据制造商推荐的方法,使用EZScan-FL for Windows程序在EZS-FL荧光板读数器(Asahi Techno Glass,Tokyo,Japan)中测量细胞内ROS产生。
2.6. 实时逆转录聚合酶链反应
在存在或不存在0.1或0.4μmol/ L萝卜硫素或5μg/ mL RAGE-Ab的情况下,用100μg/ mL的AGE-BSA或非糖基化的BSA处理周细胞4或24小时。然后,根据制造商的说明,用RNAqueous-4聚合酶链反应(PCR)试剂盒(AmbionInc,Austin,TX,USA)提取总RNA。用Prime Script RT Reagent试剂盒(Takara Bio Inc,Shiga,Japan)将总RNA逆转录成互补DNA(cDNA)。根据供应商的建议[16],使用Assay-on-Demand和TaqMan 5荧光核酸酶化学(Applied Biosystems,Foster City,CA,USA)进行定量实时PCR。简而言之,使用Premix ExTaq(Takara Bio Inc),在体积为20μL的48孔反应板中进行2ng模板cDNA的实时PCR反应。反应板在StepOne PCR系统上运行,具有以下曲线:95℃60秒,然后50个循环的95℃60秒,然后60℃15秒。在10至30个PCR循环后,通过从每个靶基因的阈值循环中减去内源基因(β肌动蛋白)的阈值的平均周期来计算每个样品中所有基因的阈值Δ周期。通过阈值表达方法的比较Δ周期来计算每个基因的相对表达。牛RAGE,MCP-1和β-肌动蛋白基因的引物和探针的ID分别为Bt03212202_m1,Bt03212321_m1和Bt03279174_g1(Applied Biosystems)。
2.7. 蛋白质印迹分析
在存在或不存在0.4μmol/ L萝卜硫素的情况下,用100μg/ mL的AGE-BSA或非糖基化的BSA处理周细胞24小时。培育后,用冷的磷酸盐缓冲盐水洗涤细胞,刮下,通过流体动力学剪切(21号针头)匀浆,并通过在12000g,4℃下离心20分钟。通过裂解缓冲液(25mmol / L 4-(2-羟乙基)-1-哌嗪乙磺酸[pH 7.1],150mmol / L氯化钠,1%Triton-X-100, 10%甘油,2mmol / L原钒酸钠,50mmol / L氟化钠,10mmol / L焦磷酸钠,1%磷酸酶抑制剂混合物I和II,和1%蛋白酶抑制剂混合物)。将样品在95℃煮沸5分钟后,通过十二烷基硫酸钠 - 聚丙烯酰胺凝胶电泳(4%-12%)分离5μg细胞裂解物,然后转移到硝酸纤维素膜上(Amersham Pharmacia Biotech,Buckinghamshire,UK)。用1:200稀释的抗RAGE多克隆抗体(Santa Cruz Biotechnology,Santa Cruz,CA,USA)或1:1000稀释的针对α微管蛋白的单克隆抗体探测膜。如前所述,用增强的化学发光检测系统(Amersham Pharmacia Biotech)使免疫反应条带可视化[25]。
2.8. 测量[3H]胸苷掺入周细胞
在存在或不存在0.1或0.4μmol/ L萝卜硫素或5μg/ mL RAGE-Ab的情况下,用100μg/ mL AGE-BSA或非糖基化BSA处理周细胞20小时。然后,加入[3H]胸苷至终浓度为1μCi/ mL,并将细胞进一步培育4小时。培育后,将细胞用冰冷的10%(wt / vol)三氯乙酸固定20分钟,然后处理所得的酸不溶性物质用于液体闪烁计数[26]。
2.9. 周细胞凋亡细胞死亡的测量
在存在或不存在0.1或0.4μmol/ L萝卜硫素或5μg/ mL RAGE-Ab的情况下,用100μg/ mL AGE-BSA或非糖基化BSA处理周细胞7天。然后,裂解细胞,并在酶联免疫吸附测定(ELISA)中分析上清液的DNA片段(Cell Death Detection ELISA plus; Roche Molecular Biochemicals,Mannheim,Germany),如前所述[2]。
2.10. 数据分析
所有值均表示为平均值±SE。进行单因素方差分析(ANOVA),然后进行Tukey或Games-Howell检验,进行统计学比较。P <0.05被认为是显著。使用由Heinrich Heine University(Dusseldorf,Germany)提供的G * Power3进行功率分析。
3. 结果
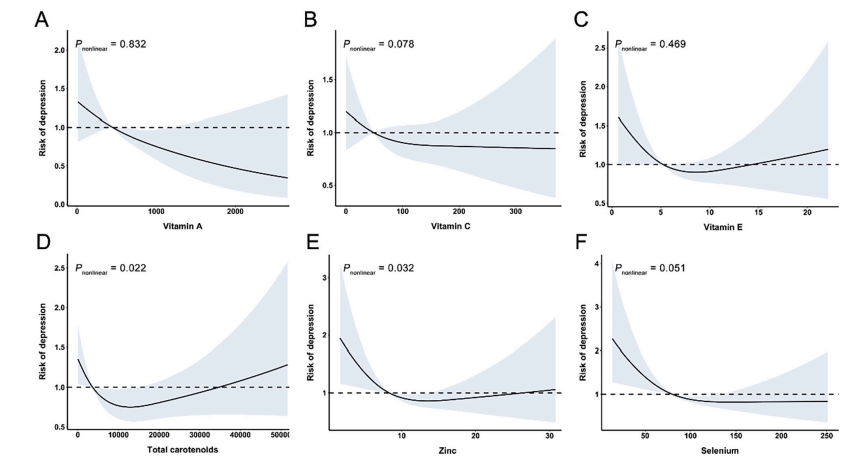
因为AGE可以通过ROS的产生对多种细胞功能发挥多效作用[5-12],我们首先研究了萝卜硫素对暴露于AGE 2小时周细胞中ROS产生的影响。如图1所示,萝卜硫素剂量依赖性地抑制AGE诱导的周细胞中ROS产生的增加; 0.4μmol/ L萝卜硫素完全阻断周细胞中AGE诱导的ROS产生。针对5μg/ mL的RAGE的抗体也完全阻断由AGE诱发的周细胞中的ROS产生。
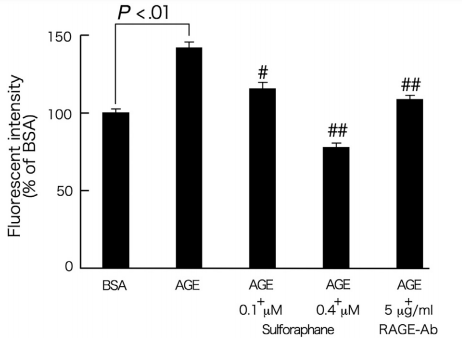
图1 - 萝卜硫素对AGE暴露的周细胞中ROS产生的影响。
在存在或不存在0.1或0.4μmol/ L萝卜硫素或5μg/ mL RAGE-Ab的情况下,用100μg/ mL AGE-BSA或非糖基化BSA处理周细胞2小时。然后,将周细胞与含有10μmol/ L 5-(和-6) - 氯甲基-2',7'-二氯二氢荧光素二乙酸酯的无酚红Dulbecco改良Eagle培养基在37℃培育15分钟。用EZS-FL荧光板读数器通过荧光强度评估细胞内ROS产生。每组n = 16。所有值均表示为平均值±SE。进行单因素方差分析,然后进行Tukey检验,进行统计学比较; P <0.05被认为是显著的。#和##,P <0.05和P <0.01分别与单独使用AGE的值相比较。 功率≥0.81。
AGE受体是AGE的信号转导受体,可以介导AGE对周细胞的有害影响[6]。通常,信使RNA(mRNA)上调可能导致蛋白质过度表达。因此,随后的研究集中于萝卜硫素对暴露于AGE 4和24小时的周细胞中RAGE基因和蛋白质表达的影响。如图2A所示,0.1和0.4μmol/ L的萝卜硫素阻止了AGE处理的周细胞中RAGE mRNA水平的上调。尽管0.1μmol/ L的萝卜硫素对RAGE蛋白表达的影响不足,但0.4μmol/ L的萝卜硫素完全阻断周细胞中AGE诱导的RAGE蛋白诱导(图2B)。
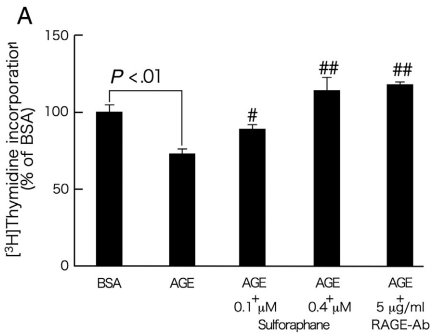
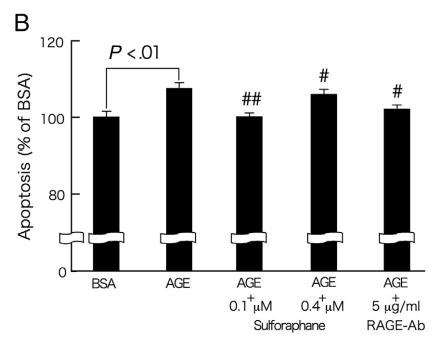
图2-萝卜硫素对AGE暴露的周细胞中RAGE基因(A)和蛋白质表达(B)的影响。
在存在或不存在0.1或0.4μmol/ L萝卜硫素的情况下,用100μg/ mL的AGE-BSA或非糖基化的BSA处理周细胞4(A)或24小时(B)。A,然后,使用Prime Script RT Reagent试剂盒将总RNA逆转录成cDNA,并使用Assay-on-Demand和TaqMan 5荧光核酸酶化学法通过实时PCR扩增。数据通过β-肌动蛋白mRNA衍生信号的强度归一化,然后与单独用BSA获得的值相关。每组n = 9。所有值均表示为平均值±SE。进行单因素方差分析,然后进行Games-Howell检验,进行统计学比较; P <0.05被认为是显著的。##,P <0.01与单独使用AGE的值比较。 功率≥0.93。B,培育后,用冷的磷酸盐缓冲盐水洗涤细胞,刮下,通过流体动力学剪切(21号针头)匀浆,并通过在12000g,4℃下离心20分钟。通过裂解缓冲液立即裂解细胞,并通过十二烷基硫酸钠 - 聚丙烯酰胺凝胶电泳(4%-12%)分离5μg细胞裂解物并转移到硝酸纤维素膜上。用1:200稀释的抗RAGE多克隆抗体或1:1000稀释的针对α微管蛋白的单克隆抗体探测膜。用增强的化学发光检测系统显现免疫反应条带。数据通过α微管蛋白衍生信号的强度归一化,然后与单独用BSA获得的值相关。每组n = 3。所有值均表示为平均值±SE。进行单因素方差分析,然后进行Tukey检验,进行统计学比较; P <0.05被认为是显著的。##,P <0.01与单独使用AGE的值比较。 功率≥0.99。
我们进一步研究了萝卜硫素对AGE暴露的周细胞中DNA合成和凋亡的影响。如图3A所示,24小时的AGE处理显著降低培养的视网膜周细胞中的DNA合成。此外,AGE暴露相对较长时间(7天)诱导周细胞的凋亡性细胞死亡(图3B)。用0.4μmol/ L萝卜硫素或5μg/ mL RAGE-Ab处理显著防止AGE的这些生长抑制和促凋亡作用(图3A和B)。此外,如图4所示,萝卜硫素剂量依赖性地抑制AGE诱导的周细胞中MCP-1 mRNA水平的上调。通过用5μg/ mL RAGE-Ab处理,通过AGE诱导4小时的周细胞MCP-1基因被完全抑制。
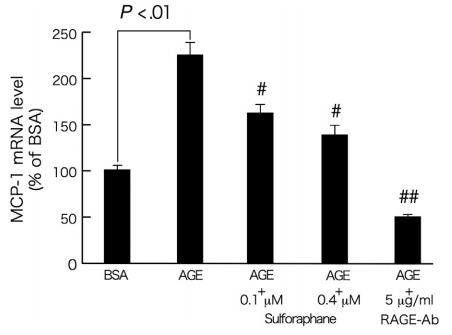
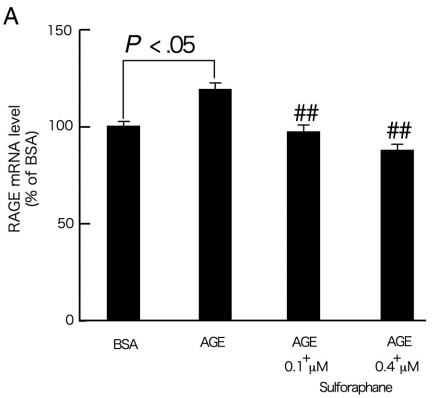
图3-萝卜硫素对AGE暴露的周细胞中[3H]胸苷掺入(A)和凋亡细胞死亡(B)的影响。
在存在或不存在0.1或0.4μmol/ L萝卜硫素或5μg/ mL RAGE-Ab的情况下,用100μg/ mL的AGE-BSA或非浓缩的BSA处理周细胞20(A)或7天(B)。A,然后,将[3H]胸苷加入培养基中。4小时后,测定[3H]胸苷掺入细胞中。每组n = 6至12。所有值均表示为平均值±SE。进行单因素方差分析,然后进行Games-Howell检验,进行统计学比较; P <0.05被认为是显著的。#和##,P <.05和P <.01分别与单独使用AGE的值相比较。 功率≥0.93。B,用细胞死亡ELISA plus检测测量凋亡细胞死亡。每组n = 18。所有值均表示为平均值±SE。进行单因素方差分析,然后进行Games-Howell检验,进行统计学比较; P <0.05被认为是显著的。#和##,P <.05和P <.01分别与单独使用AGE的值相比较。 功率≥0.81。
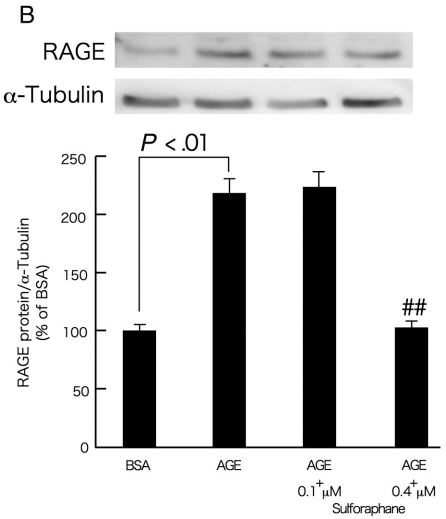
图4-萝卜硫素对AGE暴露的周细胞中MCP-1基因表达的影响。
在存在或不存在0.1或0.4μmol/ L萝卜硫素或5μg/ mL RAGE-Ab的情况下,用100μg/ mL AGE-BSA或非糖基化BSA处理周细胞24小时。然后,使用Prime Script RT Reagent试剂盒将总RNA逆转录成cDNAs,并使用Assay-on-Demand和TaqMan 5荧光核酸酶化学法通过实时PCR扩增。数据通过β-肌动蛋白mRNA衍生信号的强度归一化,然后与单独用BSA获得的值相关。每组n = 3。所有值均表示为平均值±SE。进行单因素方差分析,然后进行Tukey检验,进行统计学比较; P <.05被认为是显著的。#和##,P <0.05和P <0.01分别与单独使用AGE的值相比较。 功率≥0.79。
4. 讨论
一些大型前瞻性临床研究,糖尿病控制和并发症试验[27]以及英国前瞻性糖尿病研究[28],已经表明,强化控制血糖或血压可显著降低糖尿病患者视网膜病变发展和进展的风险。然而,严格控制血糖或血压通常难以维持,并且可能增加低血糖或低血压的风险。此外,目前治疗威胁视力的增生性糖尿病视网膜病变的治疗方案由于相当大的副作用而远远不能令人满意[5]。因此,糖尿病患者需要开发特异性靶向糖尿病性视网膜病的新型治疗策略。
微血管由2种类型的细胞,周细胞和内皮细胞组成[5,6]。以前的研究表明,周细胞有助于维持视网膜血管内稳态[4-6]。此外,在具有血小板衍生生长因子B([±]小鼠)的单一功能等位基因的小鼠中,已经显示具有周细胞的视网膜毛细血管覆盖对于内皮细胞的存活是至关重要的,特别是在诸如糖尿病的应激条件下[29]。这些观察结果表明,周细胞的丢失是糖尿病视网膜病变最早的组织病理学标志,可能使血管易于发生血管生成,血栓形成和内皮细胞损伤,从而导致糖尿病视网膜病变的全面临床表现。
该研究首次表明,萝卜硫素抑制AGE诱导的ROS产生,RAGE表达,DNA合成减少和周细胞凋亡细胞死亡。此外,RAGE-Ab完全阻止了AGE对周细胞的这些有害影响。我们之前已经证明AGE通过ROS产生刺激RAGE基因表达,并且一种抗氧化剂N-乙酰半胱氨酸可以防止AGE暴露的周细胞中DNA合成的减少以及凋亡细胞死亡[12,30,31]。目前的研究结果表明,萝卜硫素还可能抑制AGE诱导的视网膜周细胞中DNA合成和凋亡细胞死亡的减少,部分原因是通过其抗氧化特性抑制RAGE表达。此外,AGE-RAGE相互作用可诱导ROS产生并随后上调RAGE表达,这可能加强AGE对周细胞的有害作用。因此,AGE-RAGE轴和ROS之间的串扰可能随着周细胞的损伤而发生。换句话说,RAGE与AGE→ROS结合导致RAGE基因诱导,随后RAGE蛋白表达和生长抑制和炎症反应的事件将导致凋亡性细胞死亡。这是一系列可能发生在AGE暴露的周细胞中的事件。因此,我们对每次测量使用不同的处理时间。鉴于糖尿病视网膜中AGE诱发的周细胞丢失的病理作用[4-6,32,33],我们目前的研究表明,萝卜硫素可能有助于预防糖尿病视网膜病变的发展和进展。
人们越来越关注炎症反应和免疫现象在糖尿病视网膜病变发病机制中的作用[34,35]。白细胞粘附于糖尿病视网膜血管系统被认为是糖尿病视网膜病变的关键早期事件,单核细胞/巨噬细胞在糖尿病视网膜病变中积聚[34,35]。单核细胞趋化蛋白1是一种必需的趋化因子,可介导白细胞向炎性病变的募集[14]。此外,与非糖尿病患者相比,糖尿病视网膜病变中MCP-1的玻璃体水平显着升高,并且与玻璃体液中的AGE和过氧化氢水平呈正相关[36]。这些观察结果表明,萝卜硫素对周细胞中AGE诱导的MCP-1表达的抑制可能具有临床相关性。
Hanlon等[37]报道,摄入含3.9 mg萝卜硫素的液化西兰花后,萝卜硫素的血浆浓度约为70 nmol / L。 据报道十字花科蔬菜(如西兰花和卷心菜)中的萝卜硫素含量约为18至75毫克/ 100克[37,38]。因此,对本实验中使用的周细胞具有有益效果的萝卜硫素浓度(0.1-0.4μmol/ L)可与食用100g十字花科蔬菜后获得的生理水平相当。在本研究中,用0.4μmol/ L萝卜硫素处理完全阻断了AGE对ROS产生,RAGE表达和DNA合成的影响,而0.4μmol/ L萝卜硫素对细胞凋亡和MCP-1 mRNA水平的作用是部分的。晚期糖基化终产物可能导致周细胞凋亡和MCP-1基因诱导,部分以ROS非依赖性方式。尽管与体内相比,AGE的体外制备可以产生高度修饰的蛋白质,但是在体外条件下形成的非生理性AGE不太可能对周细胞产生非特异性和毒性作用。这是因为(1)体外制备的AGE的抗原表位实际上存在于糖尿病患者的血清中,并且(2)此处使用的AGE浓度(100μg/ mL)与糖尿病患者的浓度相当[39]。
在这项研究中存在一些局限性,需要额外的工作来解释ROS在萝卜硫素诱导的RAGE抑制和抑制促凋亡和炎症反应中的积极参与。在抗氧化剂如N-乙酰半胱氨酸存在下测试萝卜硫素是否也能够降低RAGE表达并抑制周细胞中AGE的有害作用将是有趣的。此外,由于AGE和萝卜硫素对细胞凋亡的影响是适度的,末端脱氧核苷酸转移酶介导的2'-脱氧尿苷5'-三磷酸缺口末端标记染色将是有用的。因为mRNA水平并不总是与蛋白质表达相关,所以检查萝卜硫素是否实际上可以减少AGE暴露的周细胞产生MCP-1也是有帮助的。此外,通过二氢乙锭图像染色评估萝卜硫素对ROS产生的影响将加强目前的发现。虽然我们的体外研究结果表明,食用100克十字花科蔬菜可以防止周细胞损伤,但需要进一步的前瞻性研究来阐明摄入的萝卜硫素的量是否有助于预防糖尿病患者的视网膜病变。
我们目前的研究和其他研究表明,萝卜硫素可以抑制AGE暴露的周细胞中DNA合成,细胞凋亡和炎症反应的减少,部分是通过其抗氧化特性抑制RAGE。鉴于萝卜硫素的浓度对本实验中使用的周细胞具有有益作用,它可以与食用100g十字花科蔬菜后获得的生理水平相当。通过萝卜硫素封闭周细胞中的AGE-RAGE轴可能是治疗糖尿病性视网膜病变的新型治疗靶点,值得进一步研究。
本文由福山生物整理翻译,转载请注明出处。